2 TINJAUAN PUSTAKA - repository.ipb.ac.id · daun yang menonjol serupa sendi-sendi tulang. Susunan...
22
2 TINJAUAN PUSTAKA Deskripsi Jenis Api-api (A. marina (Forsk.) Vierh. 1907) Api-api adalah nama sekelompok tumbuhan dari marga Avicennia, suku Acanthaceae (Wikipedia 2007). Dalam sistem klasifikasi, tanaman A. marina mempunyai penggolongan sebagai berikut (Plantamor 2012): Kingdom Divisi Kelas Ordo Family Genus Jenis : : : : : : : Plantae Magnoliophyta Magnoliopsida Scrophulariales Acanthaceae Avicennia A. marina (Forsk.) Vierh. Nama lokal : Api-api jambu, sia-sia putih, api-api, pejapi, nyapi, api, sia, hajusa, pai. (Kusmana et al. 2008). Api-api biasa tumbuh di tepi atau dekat laut sebagai bagian dari komunitas hutan bakau. Nama Avicennia dilekatkan pada genus ini untuk menghormati Ibnu Sina, di dunia barat terkenal sebagai Avicenna, salah seorang pakar dan perintis kedokteran modern dari Persia (Wikipedia 2007). Api-api merupakan salah satu jenis yang termasuk ke dalam kelompok mangrove utama. Adapun karakteristik mangrove utama sebagai berikut (Kusmana et al. 2008): a. Hanya hidup di habitat mangrove, tidak dapat tumbuh menyebar ke daratan. b. Berperan penting dalam struktur komunitas mangrove dan mampu membentuk tegakan murni. c. Memiliki morfologi spesifik sebagai hasil adaptasi terhadap lingkungan, seperti adanya akar permukaan (akar napas/akar udara) dan buah vivipar. Sebagai warga komunitas mangrove, api-api memiliki beberapa ciri yang merupakan bagian dari adaptasi pada lingkungan berlumpur dan bergaram, diantaranya akar nafas (pneumatophores) yang muncul 10-30 cm dari substrat, seperti paku dengan diameter 0.5-1 cm. Akar nafas api-api yang padat, rapat dan banyak sangat efektif untuk menangkap dan menahan lumpur sehingga mempercepat proses pembentukan tanah timbul serta berbagai sampah yang terhanyut di perairan. Jalinan perakaran ini juga menjadi tempat mencari makanan bagi aneka jenis kepiting bakau, siput dan teritip (Wikipedia 2007; Kusmana et al. 2005b). d. Secara fisiologis memiliki mekanisme untuk mengeluarkan garam dari tubuhnya. Api-api memiliki daun dengan kelenjar garam. Daun api-api berwarna putih sampai keabu-abuan dilapisi kristal garam di sisi bawahnya. Ini adalah kelebihan garam yang dibuang oleh tumbuhan tersebut (Wikipedia 2007; Kusmana et al. 2008). e. Relatif terisolasi secara taksonomi dari komunitas daratan, minimal pada level marga (genus). Api-api menyukai rawa-rawa mangrove, tepi pantai yang berlumpur, atau di sepanjang tepian sungai pasang surut. Beberapa jenisnya seperti A. marina
-
Upload
truongdang -
Category
Documents
-
view
242 -
download
0
Transcript of 2 TINJAUAN PUSTAKA - repository.ipb.ac.id · daun yang menonjol serupa sendi-sendi tulang. Susunan...

2 TINJAUAN PUSTAKA
Deskripsi Jenis Api-api (A. marina (Forsk.) Vierh. 1907)
Api-api adalah nama sekelompok tumbuhan dari marga Avicennia, suku
Acanthaceae (Wikipedia 2007). Dalam sistem klasifikasi, tanaman A. marina
mempunyai penggolongan sebagai berikut (Plantamor 2012):
Kingdom
Divisi
Kelas
Ordo
Family
Genus
Jenis
:
:
:
:
:
:
:
Plantae
Magnoliophyta
Magnoliopsida
Scrophulariales
Acanthaceae
Avicennia
A. marina (Forsk.) Vierh.
Nama lokal
: Api-api jambu, sia-sia putih, api-api, pejapi, nyapi, api, sia,
hajusa, pai. (Kusmana et al. 2008).
Api-api biasa tumbuh di tepi atau dekat laut sebagai bagian dari komunitas
hutan bakau. Nama Avicennia dilekatkan pada genus ini untuk menghormati Ibnu
Sina, di dunia barat terkenal sebagai Avicenna, salah seorang pakar dan perintis
kedokteran modern dari Persia (Wikipedia 2007).
Api-api merupakan salah satu jenis yang termasuk ke dalam kelompok
mangrove utama. Adapun karakteristik mangrove utama sebagai berikut
(Kusmana et al. 2008):
a. Hanya hidup di habitat mangrove, tidak dapat tumbuh menyebar ke daratan.
b. Berperan penting dalam struktur komunitas mangrove dan mampu membentuk
tegakan murni.
c. Memiliki morfologi spesifik sebagai hasil adaptasi terhadap lingkungan, seperti
adanya akar permukaan (akar napas/akar udara) dan buah vivipar.
Sebagai warga komunitas mangrove, api-api memiliki beberapa ciri yang
merupakan bagian dari adaptasi pada lingkungan berlumpur dan bergaram,
diantaranya akar nafas (pneumatophores) yang muncul 10-30 cm dari substrat,
seperti paku dengan diameter 0.5-1 cm. Akar nafas api-api yang padat, rapat
dan banyak sangat efektif untuk menangkap dan menahan lumpur sehingga
mempercepat proses pembentukan tanah timbul serta berbagai sampah yang
terhanyut di perairan. Jalinan perakaran ini juga menjadi tempat mencari
makanan bagi aneka jenis kepiting bakau, siput dan teritip (Wikipedia 2007;
Kusmana et al. 2005b).
d. Secara fisiologis memiliki mekanisme untuk mengeluarkan garam dari
tubuhnya.
Api-api memiliki daun dengan kelenjar garam. Daun api-api berwarna
putih sampai keabu-abuan dilapisi kristal garam di sisi bawahnya. Ini adalah
kelebihan garam yang dibuang oleh tumbuhan tersebut (Wikipedia 2007;
Kusmana et al. 2008).
e. Relatif terisolasi secara taksonomi dari komunitas daratan, minimal pada level
marga (genus).
Api-api menyukai rawa-rawa mangrove, tepi pantai yang berlumpur, atau
di sepanjang tepian sungai pasang surut. Beberapa jenisnya seperti A. marina

5
(Gambar 1) memperlihatkan toleransi yang tinggi terhadap kisaran salinitas,
mampu tumbuh di rawa air tawar hingga di substrat yang berkadar garam sangat
tinggi. Kebanyakan jenisnya merupakan jenis pionir dan oportunistik, serta mudah
tumbuh kembali. Pohon-pohon api-api yang tumbang atau rusak dapat segera
trubus (bersemi kembali), sehingga mempercepat pemulihan tegakan yang rusak
(Wikipedia 2007; Kusmana et al. 2008).
A. marina memiliki ukuran pohon kecil atau besar, tinggi mencapai 30 m,
dengan tajuk yang agak renggang. Pepagan (kulit batang) halus keputihan sampai
dengan abu-abu kecoklatan dan retak-retak. Ranting memiliki buku-buku bekas
daun yang menonjol serupa sendi-sendi tulang. Susunan daun tunggal berhadapan
dengan helaian berbentuk elips dan ujung daun akut sampai membundar
berukuran panjang 5-11 cm. Api-api memiliki biji kriptovivipar. Bunga muncul
terutama pada bulan juli-februari, sedangkan munculnya buah pada bulan
november-maret (musim hujan), dengan antesis sampai kemasakan 2-3 bulan.
Bunga bersifat infloresensi berjumlah 8-14, dengan bulir rapat, panjang mencapai
1-2 cm, dengan susunan terminal atau aksilar pada tunas-tunas distal dengan daun
mahkota berjumlah 4, berwarna kuning sampai oranye. Kelopak memiliki 5
cuping dan benang sari sebanyak 4 buah berukuran 0.4-0.5 cm. Lebar buah 1.5-
2.0 cm dan panjang 1.5-2.5 cm dengan perikarp berwarna hijau, bagian dalam
hijau sampai coklat muda/kekuningan dan pada permukaan terdapat rambut halus.
Buah membundar secara apikal atau dengan sebuah paruh yang pendek
(Wikipedia 2007; Kusmana et al. 2008).
Gambar 1 Avicennia marina (Forsk.) Vierh. 1907.

6
Deskripsi Jenis Bakau (R. mucronata Lamk. 1804)
Dalam sistem klasifikasi, tanaman R. mucronata mempunyai
penggolongan sebagai berikut:
Kingdom
Divisi
Kelas
Ordo
Family
Genus
Jenis
:
:
:
:
:
:
:
Plantae
Magnoliophyta
Magnoliopsida
Malpighiales
Rhizophoraceae
Rhizophora
R. mucronata Lamk.
Nama lokal
: bakau, bako-gandul, bakau-genjah, bakau-bandul, bakau-
hitam, tanjang-lanang, tokke-tokke, bakao, bakau-laki,
blukap, tongke-besar, lului, bakau-bakau, wako, bako,
bangko, blukap (Kusmana et al. 2008).
R. mucronata (Gambar 2) merupakan jenis mangrove utama dengan tinggi
batang mencapai 27 m, jarang melebihi 30 m. Umumnya tumbuh di zona terluar,
mengembangkan akar tunjang (stilt root) untuk bertahan dari ganasnya
gelombang. R. mucronata memiliki akar tunjang yang besar dan berkayu dan akar
udara yang tumbuh dari percabangan bagian bawah. Batang memiliki diameter
hingga 70 cm dengan kulit kayu berwarna gelap hingga hitam dan terdapat celah
horizontal/memecah datar. Daun tunggal berhadapan dengan gagang daun
berwarna hijau, berbentuk elips melebar hingga bulat memanjang dengan ujung
daun berarista (aristate) (ujung daun mirip gigi yang meramping tajam). Panjang
daun mencapai 15-20 cm, lebih besar dari R. stylosa, dengan bagian paling lebar
berada di tengah. Permukaan bawah daun hijau kekuningan dan terdapat bintik-
bintik hitam kecil yang tersebar. Pinak daun terletak pada pangkal gagang daun
berukuran 5.5-8.5 cm. (Noor et al. 1999; Kusmana et al. 2008).
R. mucronata memiliki biji vivivar dan bunga infloresensi, bercabang-
cabang melalui pembagian menjadi dua secara berulang kali (dichotomous),
berbunga sebanyak 4-8 dengan perbungaan terbatas (cyme), menggantung, dan
aksilar. Daun mahkota berjumlah 4, berwarna putih, dan berambut dengan
kelopak bercuping 4, berwarna kuning keputihan sampai hijau kekuningan.
Benang sari berjumlah 8 dengan diameter 3-4 cm dan panjang 1.5-2.0 cm.
Tangkai putik pendek dengan kepala putik hampir duduk (hampir tanpa tangkai).
Buah berdiameter 2.0-2.3 cm, sedangkan panjang 50-70 cm berwarna hijau
sampai hijau kekuningan, leher kotiledon kuning ketika masak, dengan
permukaan berkutil (mempunyai struktur mirip kutil). R. mucronata berbuah
silindris (hipokotil), rontok dari bawah leher kotiledon, mengapung, dan tersebar
oleh arus. Pemunculan bunga sepanjang tahun (terutama agustus-desember) dan
pemuculan buah pada bulan oktober-desember (awal musim hujan), dengan
antesis sampai kemasakan sekitar 14-15 bulan (Kusmana et al. 2008).
R. mucronata tumbuh di tepi sungai-sungai kecil, pantai yang berawa dan
berlumpur tanpa ada ombak yang kuat, dan tumbuh baik di wilayah sungai
estuaria dengan lumpur mangrove yang lunak. Jarang sekali tumbuh pada daerah
yang jauh dari air pasang surut. Pertumbuhan optimal terjadi pada areal yang
tergenang dalam, sedikit kandungan pasirnya, serta pada tanah yang kaya akan
humus. R. mucronata teradaptasi dengan berbagai elevasi dengan kisaran yang

7
lebar. Jenis ini lebih toleran terhadap substrat yang lebih keras dan berpasir bila
dibandingkan dengan jenis R. apiculata. menyebar luas mulai dari Afrika timur,
Madagaskar, Mauritania, Asia Tenggara, kepulauan Nusantara, Melanesia dan
Mikronesia. Pada saat ini telah diintroduksikan ke daerah Hawaii (Noor et al.
1999; Kusmana et al. 2005b; Kusmana et al. 2008).
Gambar 2 R. mucronata Lamk. 1804.
Teknik Rehabilitasi Mangrove
1. Penanaman dengan propagul (Kusmana et al. 2009a)
Penanaman langsung dengan menggunakan propagul umumnya dilakukan
apabila areal penanaman berupa tanah lumpur. Penanaman propagul ini dilakukan
dengan cara membenamkan seperempat sampai sepertiga panjang propagul ke
dalam lumpur secara tegak dengan bakal kecambah menghadap ke atas. Jika
propagul ditanam terlalu dalam, lumpur akan menutup lentisel, dan hipokotil tidak
dapat berespirasi, dan hal ini akhirnya dapat menyebabkan kematian. Demikian
juga sebaliknya, apabila propagul ditanam terlalu dangkal, dia akan mudah hanyut
oleh ombak dan air pasang. Untuk R. mucronata, R. apiculata dan R. stylosa,
kelopak buah (calyx) harus selalu dilepas sebelum penanaman (biasanya kalau
propagul sudah matang, calyx ini akan lepas dengan sendirinya bersama perikarp).
Di lain pihak, untuk B. gymnorrhiza, kelopak buah tersebut harus tetap dibiarkan
utuh ketika penanaman. Calyx pada B. gymnorrhiza akan rontok sendiri setelah
seminggu. Bila setelah seminggu calyx belum rontok, calyx ini perlu dilepas
dengan tangan, tapi tidak boleh dengan cara paksa. Apabila area penanaman
terdiri atas tanah lumpur yang kurang lembek, penanaman propagul dilakukan

8
pada lubang tanam yang dibuat dengan tugal (galah kayu yang ujungnya
diruncingkan).
2. Penanaman dengan bibit (Kusmana et al. 2009a)
Bibit ditanam pada lobang tanam yang ukurannya sebesar ukuran polibag
media bibit (Gambar 3). Penanaman bibit mangrove di lahan pinggir sungai,
pinggir pantai dan daerah-daerah lainnya dengan arus air yang relatif kuat
disarankan polibagnya tidak disobek. Adapun penanaman bibit di lahan-lahan
yang arus pasang surutnya relatif tenang polibag disarankan dirobek dengan cara
disayat secara hati-hati sebelum dimasukkan ke lubang tanam. Polibag bekas
tersebut kemudian disangkutkan di ujung ajir sebagai tanda bahwa anakan sudah
ditanam. Kemudian, tanah atau lumpur ditimbunkan kedalam lubang tanam
sehingga propagul dapat berdiri tegak. Kemudian bila perlu, propagul, tersebut
diikatkan pada ajir, supaya tanaman kokoh kedudukannya dan tidak mudah
terbawa arus air. Yang perlu diperhatikan bila tanaman diikatkan ke ajir adalah
bahwa ajir itu sendiri harus kokoh kedudukannya di substrat mangrove (dalam hal
ini misalnya, ajir cukup dalam ditancapkannya ke lumpur mangrove). Bila
kedudukan (penjangkaran ke substrat mangrove) ajir lebih lemah dibanding bahan
tanaman, maka pengikatan tanaman ke tiang ajir, malah membebani tanaman dan
malah memperbesar peluang hanyutnya tanaman oleh arus.
Jika terjadi penundaan penanaman di lokasi penanaman, padahal bahan
tanaman sudah diangkut ke lokasi, bahan-bahan tanaman tersebut sebaiknya
disimpan di tempat yang teduh. Bahan tanaman berupa propagul sebaiknya
disimpan dalam posisi tegak di areal yang berlumpur, dan teduh.
Gambar 3 Penananaman anakan ke dalam lubang tanam.
3. Sistem tanam (Kusmana et al. 2009a)
Ada dua sistem penanaman mangrove yang umum dilakukan, yakni,
sistem banjar harian (penanaman seluruh areal) dan sistem tumpang sari
(wanawina/silvofishery). Secara umum tidak terdapat perbedaan secara prinsip
dalam cara penanaman dari kedua sistem tersebut. Khusus pada sistem tumpang
sari, terdapat tambahan kegiatan dalam tahapan persiapan lapangan, yakni
pembuatan konstruksi tambak, saluran air dan tapak tanam seperti terlihat pada
Gambar 4.
polibag
Lubang tanam
tanah

9
Gambar 4 Model sistem wanamina yang umum di Indonesia.
4. Teknik rehabilitasi pada tapak-tapak khusus
a. Tapak berarus dan berombak besar (Kusmana et al. 2009b)
Areal penanaman mangrove pada tapak berarus dan berombak besar
umumnya terdapat pada tepi laut lepas atau daerah cekungan tepi laut dengan
pusaran arus deras dan gelombang besar. Sebelum dilakukan penanaman terlebih
dahulu dibuat penahan arus dan pemecah gelombang (water break) di depan lahan
yang akan ditanami. Bentuk-bentuk penahan arus dan pemecah gelombang dapat
berupa: (a) tumpukan batu yang dimasukkan ke dalam anyaman kawat
(beronjong), (b) berupa tripod (cetak beton berkaki tiga), (c) gundukan atau
guludan tanah/batu (rubble mould), dan (d) anyaman cerucuk bambu/kayu.
Bentuk-bentuk penahan arus dan pemecah ombak (water break) dalam
penanaman mangrove pada tapak berarus deras berombak besar dapat dilihat pada
Gambar 5. Penahan arus dan pemecah gelombang bentuk gundukan batu (rubble
mould) dapat dilihat pada Gambar 6.
SALURAN AIR
PINTU AIR
LA
HA
N T
EM
PA
TM
EM
EL
IHA
RA
IK
AN
LA
HA
N T
EM
PA
T M
EM
ELIH
AR
A IK
AN
LA
HA
N T
EM
PA
T M
EM
ELIH
AR
A IK
AN
LAHAN TEMPAT MEMELIHARA IKAN
LAHAN TEMPAT MEMELIHARA IKAN
PINTU AIR
SALURAN AIR
tegakan
mangrove
ko
lam
pintu air saluran air
tanggul
ko
lam
tegakan
mangrove
tanggul
saluran air Pintu air
ko
lam
tegakan
mangrove
tanggul
saluran air pintu air

10
SEA
DITCH
PLANTING AREA
SEA
DITCH
PLANTING AREA
TRIPOD
TRIPOD
TRIPO
D
SEA
DITCH
PLANTING AREA
STONE DEPOSITION
PLANTING AREA
SEA
BAMBOO STICK
DITCH
Gambar 5 Pemecah ombak berupa tumpukan batu yang dimasukkan ke dalam
kawat (a), berupa tripod (b) guludan tanah (c) dan cerucuk bambu
dan kayu (d).
Gambar 6 Penahan arus dan pemecah gelombang bentuk gundukan batu (rubble
mould).
(a) (b)
laut laut
Area penanaman Area penanaman
(d) (c)
laut laut
Area penanaman Area penanaman

11
Untuk tapak semacam ini, sebaiknya digunakan bibit jenis Rhizophora
spp., terutama R. mucronata. Jarak tanam sebaiknya cukup rapat (misal 1 x 1 m
atau lebih rapat) dengan berselang seling, sehingga membentuk pola “untu
walang” (zig zag).
Agar anakan yang ditanam tidak mudah hanyut, maka sebaiknya anakan
tersebut diikatkan pada tiang pancang/bambu (Gambar 7).
1. Penggunaan tiang pancang
Tiang pancang yang terbuat dari kayu atau bambu (diameter minimal 7.5
cm, panjang 1 m, dan runcing di bagian bawahnya) ditancapkan ke dalam lumpur
sedalam 0.5 m, tepat di samping semai mangrove yang ditanam. Batang semai
tanaman diikatkan pada tiang pancang. Untuk memperoleh kedudukan yang lebih
kuat, ruas bambu tiang tersebut dilubangi terlebih dahulu, kemudian lumpur
dimasukkan ke dalam tiangnya saat tiang ditancapkan.
2. Penggunaan ruas bambu besar
Bambu yang diameter 20 – 25 cm dan tinggi 1 m, ditancapkan ke dalam
lumpur sedalam 0.5 m pada lokasi dimana semai mangrove akan ditanam. Bambu
dilubangi ruas dalamnya dan diperuncing pada bagian bawahnya. Isilah bambu
dengan lumpur, kemudian tanamlah semai mangrove ke dalam bambu tersebut.
Salah satu jenis bambu yang berukuran sebesar itu adalah bambu betung
(Dendrocalamus asper).
Gambar 7 Penguat tanaman di tapak yang berombak besar menggunakan tiang
pancang (a) dan menggunakan bambu besar (b).
b. Tapak dengan arus deras pinggir sungai (Kusmana et al. 2009b)
Penanaman mangrove pada tapak dengan arus deras pinggir sungai
dilakukan dengan menggunakan jarak tanam atau tanpa menggunakan jarak
tanam. Jika menggunakan jarak tanam sebaiknya digunakan jarak tanam rapat
kurang dari 0.5 m x 0.5 m.
Pola tanam bisa menggunakan model zig-zag (untu walang). Penanaman
tanpa menggunakan jarak tanam sering disebut dengan penanaman dengan teknik
gerombol (sistem cluster). Mengingat arus air sungai yang deras maka penanaman
mangrove pada tapak berarus deras tepi sungai ini mutlak diperlukan ajir untuk
mengikat tanaman agar tidak terbawa arus.
Ajir bisa berupa bambu atau kayu. Bibit tanaman yang di tanam
selanjutnya dengan menggunakan tali rafia diikat dengan ajir bambu atau kayu
tersebut. Untuk menghindari hanyutnya media tanah yang terdapat dalam polibag

12
0,25 – 0.5 m
0,5-1 m
0,5 - 1 m
oleh arus sungai yang deras sebaiknya pada waktu penanaman polibag tidak perlu
dibuka, cukup diperbanyak lobang-lobang akar pada polibagnya. Sketsa pola
penanaman mangrove pada tapak berarus deras tepi sungai dapat dilihat pada
Gambar 8.
Gambar 8 Sketsa pola penanaman mangrove pada tapak berarus deras tepi sungai
dengan pola zig-zag (untu walang).
c. Tapak berlumpur dalam (Kusmana et al. 2009b)
Tapak berlumpur dalam bisa terdapat pada areal penanaman mangrove tepi
laut, tepi sungai atau bekas tambak. Pada tapak yang berlumpur dalam, sebaiknya
digunakan bibit atau propagul R. mucronata. Seperti halnya pada tapak yang
berombak besar, bibit atau propagul mangrove yang ditanam diikatkan pada tiang
pancang. Alternatif lain bibit yang akan ditanam dimasukkan ke dalam bambu
yang telah berisi media tanah. Jarak tanam yang dipakai sebaiknya jarak tanam
rapat (maksimal 1 x 1 m).
d. Tapak berbatu atau berkerikil (Kusmana et al. 2009b)
Tapak berbatu atau berkerikil umumnya ditemukan pada areal penanaman
mangrove di dekat terumbu karang atau di pantai-pantai terjal berdinding batu
atau berkerikil. Prinsip penanaman mangrove pada tapak berbatu atau berkerikil
ini adalah memindahkan batu atau berkerikil yang terdapat pada lobang tanam
dengan media lumpur atau tanah.
Teknik penanaman mangrove pada tapak berbatu/berkerikil dapat
menggunakan teknik jarak tanam rapat atau tanpa menggunakan jarak tanam
(penanaman bergerombol/cluster). Penanaman dengan jarak tanam dapat
menggunakan bibit dengan lubang tanam yang besar dan diganti dengan lumpur.
Penanaman dengan gerombol/cluster disesuaikan dengan sebaran dan ketebalan
batu/kerikil yang ada. Dalam satu titik penanaman bisa ditanam lebih dari satu
bibit mangrove. Jika arus/gelombang tidak besar tidak diperlukan ajir tanaman.
Teknik penanaman gerombol/cluster pada tapak berbatu/berkerikil dapat dilihat
pada Gambar 9 dan 10.

13
Gambar 9 Teknik penanaman mangrove pada tapak berbatu/berkerikil dengan
cara gerombol (cluster).
Gambar 10 Teknik penanaman pada tapak berbatu/berkerikil dengan lubang besar
dan diberi lumpur.
e. Tapak tertimbun pasir pasca tsunami (Kusmana et al. 2009b)
Tapak tertimbun pasir terjadi akibat gelombang laut yang besar atau
tsunami. Pasca terjadinya tsunami selain menghancurkan berbagai sarana
prasarana di tepi pantai juga sering menyisakan timbunan pasir yang luas dan
tebal. Dalam rangka rehabilitasi dan penanaman mangrove di kawasan ini
diperlukan usaha mengurangi timbunan pasir sebelum penanaman.
Puslitbang Hutan dan Konservasi Alam, Badan Penelitian dan
Pengembangan Kehutanan telah mencoba menanam mangrove pada areal yang
tertimbun pasir pasca tsunami di Aceh dengan cara menggunakan polybag
berukuran besar, pembuatan parit, dan lubang tanam berukuran besar yang diisi
dengan lumpur. Walaupun pengaruhnya terhadap pertumbuhan anakan belum
diperoleh, namun ada indikasi anakan mangrove dapat tumbuh secara baik dengan
perlakuan tersebut.
polybag
Lubang tanam yang lebar dan dalam
diisi lumpur
pasir

14
0.5 – 0.6 m
pasir
parit atau lubang yang diisi
dengan lumpur bibit mangrove
Polibag berukuran besar
pasir
Prinsip yang dipakai dalam penanaman mangrove pada tapak tertimbun
pasir sama halnya dengan tapak berbatu berkerikil yaitu menggali, memindahkan
dan mengganti pasir yang ada di lubang tanaman dengan lumpur. Bentuk-bentuk
penanaman pada tapak yang tertimbun pasir dapat dilihat pada Gambar 11 dan
Gambar 12.
Gambar 11 Teknik penanaman mangrove pada tapak tertimbun pasir dengan
mengganti lubang tanam dengan lumpur atau menggunakan polybag
berukuran besar.
Gambar 12 Teknik penanaman mangrove tertimbun pasir dengan cara penggalian
parit-parit yang diisi lumpur.
f. Tapak dengan air tergenang dalam dan diam (Kusmana et al. 2009b)
Tapak tanaman mangrove pada air tergenang dalam dan diam (tidak
berarus deras) umumnya terdapat pada kawasan hutan mangrove yang mengalami
degradasi seperti bekas tambak, bekas galian atau bekas saluran. Kedalaman air
bervariasi yang umumnya lebih dari 1.5 m sampai 3 m. Lokasi bekas galian
tersebut dapat ditemukan di dekat pantai yang terkena pasang-surut harian atau
jauh dari pantai yang tidak tidak terjangkau oleh pasang surut pantai sehingga
tingkat salinitas air genangan bervariasi.
Teknik rehabilitasi pada tapak dengan air tergenang dalam dan tidak
berarus deras ini dengan menggunanakan sistem guludan bambu. Teknik guludan
bambu ini dikembangkan oleh Kusmana et al. (2005a) untuk merehablitasi
mangrove tergenang air dalam di sekitar Tol Sedyatmo, wilayah Jakarta Utara.
Hasil penanaman mangrove dengan teknik guludan bambu tersebut berhasil

15
dengan baik. Selanjutnya teknik tersebut dikembangkan untuk merehabilitasi
kawasan mangrove yang tergenang air dalam di beberapa lokasi di Jakarta.
Prinsip dasar yang digunakan dalam sistem guludan bambu tersebut adalah
memperpendek genangan air sampai pada zona perakaran bibit mangrove.
Guludan dibuat dari cerucuk bambu yang dipasang rapat seperti pagar berbentuk
persegi panjang. Cerucuk bambu tersebut diikat dengan bambu penjepit di bagian
atas dan bawah. Pagar cerucuk bambu tersebut selanjutnya diisi karung goni berisi
tanah urugan. Tumpukan karung dalam cerucuk bambu dibuat sampai 20 cm di
bawah permukaan air. Selanjutnya tumpukan karung tersebut ditimbun dengan
tanah curah yang berisi lumpur sampai kira-kira 20 cm di atas permukaan air
(Gambar 13). Setelah proses stabilitasi tanah dapat dilakukan pemasangan ajir dan
penanaman bibit tanaman mangrove. Jarak tanam yang digunakan sebaiknya jarak
tanam rapat kurang dari 1 x 1 m.
Gambar 13 Struktur guludan (Kusmana 2010).
Model Pertumbuhan
Model adalah contoh sederhana yang mewakili atau menggambarkan suatu
sistem yang nyata. Model itu sendiri dibangun dari hasil penelitian atau
pengalaman yang berulang-ulang, sehingga tercipta suatu pengetahuan. Oleh
karena itu, model memiliki peranan penting di dalam ilmu pengetahuan.
Penyusunan model sangat penting dalam suatu penelitian, terutama untuk
menghemat waktu dan biaya (Harja dan Rahayu 2010).
Siswadi (1991) mengemukakan bahwa suatu model seringkali
dikelompokkan antara lain berdasarkan (a) upaya memperolehnya, (b) keterkaitan
pada waktu, atau (c) sifat keluarannya. Model yang berdasarkan upaya
memperolehnya misalnya adalah: model teoritik, mekanistik, dan empirik. Model
teoritik digunakan sebagai model yang diperoleh dengan menggunakan teori-teori
yang berlaku. Model mekanistik digunakan bila model tersebut diperoleh
berdasarkan mekanisme pembangkit fenomena. Model empirik digunakan bagi
model yang diperoleh hanya dari pengamatan tanpa menjelaskan sama sekali
tentang mekanismenya. Model yang didasarkan keterkaitannya pada waktu adalah
model statik dan dinamik. Model statik adalah model yang tidak terkait dengan
waktu, sedangkan model dinamik tergantung pada waktu. Bila perubahan dalam
model dinamik terjadi atau diamati secara kontinyu dalam waktu, maka model

16
tersebut dikatakan sebagai model kontinyu, bila tidak, maka model tersebut
dikatakan sebagai model diskret.
Handoko (2005) mengelompokkan beberapa model sebagai berikut:
1. Model empirik dan mekanistik
Model empirik dibuat berdasarkan pengamatan empirik/statistik, tanpa
menjelaskan atau didasarkan atas proses terjadinya. Model mekanistik
menjelaskan mekanisme proses terjadinya dalam suatu sistem.
2. Model deskriptif dan model numerik
Model deskriptif menggambarkan bentuk-bentuk hubungan secara
konsepsi atau berupa simbol-simbol tanpa mengandung bentuk hubungan
numerik. Model numerik menggambarkan hubungan-hubungan dalam bentuk
persamaan-persamaan matematik.
3. Model dinamik dan statik
Model dinamik menjelaskan tentang unsur waktu sebagai peubah penting.
Model statik tidak menjelaskan peubah-peubah yang ada sebagai fungsi waktu.
4. Model deterministik dan stokastik
Model deterministik tidak memperhitungkan peluang terjadinya kesalahan
hasil prediksi. Model stokastik merupakan suatu model dengan hasil prediksi yang
mengandung toleransi yang dapat berupa simpangan yang secara statistik dapat
digambarkan dengan ragam, simpangan baku, dan koefisien keragaman.
Pertumbuhan tanaman merupakan sistem yang dinamik, sehingga model
dinamik merupakan model yang sesuai terhadap pertumbuhan tanaman. Menurut
Davis dan Jhonson (1987) pertumbuhan didefinisikan sebagai pertambahan dari
jumlah dan dimensi pohon, baik diameter maupun tinggi yang terdapat pada suatu
tegakan. Pertumbuhan ke atas (tinggi) merupakan pertumbuhan primer (initial
growth), sedangkan pertumbuhan ke samping (diameter) disebut pertumbuhan
sekunder (secondary growth).
Diameter merupakan salah satu dimensi pohon yang paling sering
digunakan sebagai parameter pertumbuhan. Pertumbuhan diameter dipengaruhi
oleh faktor-faktor yang mempengaruhi fotosintesis. Pertumbuhan diameter
berlangsung apabila keperluan hasil fotosintesis untuk respirasi, penggantian
daun, pertumbuhan akar, dan tinggi telah terpenuhi (Davis dan Jhonson 1987).
Menurut Sitompul dan Guritno (1995), model pertumbuhan biasanya
berkenaan dengan hubungan diantara proses pertumbuhan (yang dinyatakan
dalam produknya) dengan faktor pengendali utama produknya dalam bentuk
persamaan. Kebanyakan model pertumbuhan pada masa lampau bersifat empiris
yaitu fungsi kadang-kadang dipilih dengan melihat data begitu saja dan membuat
suatu penaksiran karena tujuannya, biasanya hanya untuk mendapatkan suatu
ringkasan matematik dari data mengenai pertumbuhan keseluruhan tanaman atau
bagian tanaman, sehingga parameter model sering kurang atau tidak mempunyai
arti biologi. Akan tetapi, usaha belakangan ini telah mencoba memilih fungsi yang
logis secara biologi dengan parameter-parameter yang dapat menggambarkan
sesuatu mekanisme fisiologi atau biokimiawi yang mendasari proses
pertumbuhan. Bentuk pertumbuhan berubah menjadi asimptotis jika substrat
pertumbuhan seperti fotosintat atau unsur hara menjadi terbatas atau menurun
dengan adanya proses penuaan atau senesens.
Pola pertumbuhan tegakan antara lain dinyatakan dalam bentuk kurva
pertumbuhan yang merupakan hubungan fungsional antara sifat tertentu tegakan

17
antara lain volume, tinggi, bidang dasar, dan diameter dengan umur tegakan.
Bentuk kurva pertumbuhan tegakan yang ideal akan mengikuti bentuk ideal bagi
pertumbuhan organisme, yaitu bentuk sigmoid. Bentuk umum kurva pertumbuhan
kumulatif tumbuh-tumbuhan akan memiliki tiga tahap, yaitu tahap pertumbuhan
eksponensial, tahap pertumbuhan mendekati linear, dan pertumbuhan asimptotis
(Davis dan Jhonson 1987).
Menurut Fekedulegn et al. (1999), berbagai model pertumbuhan yang
umumnya digunakan dalam bidang kehutanan tertera pada Tabel 1.
Tabel 1 Model pertumbuhan tanaman (Fekedulegn et al. 1999)
Model Bentuk persamaan Sumber
Negatif
eksponensial f(t) = a(1-exp(-kt))+e Philip (1994)
Monomolekular f(t) = a(1-b exp(-kt))+e Draper dan Smith (1981)
Mitcherlich f(t) = (a-bkt)+e Philips dan Campbell
(1968)
Gompertz f(t) = a exp(-b exp(-kt))+e Draper dan Smith (1981)
Logistik f(t) = a/(1+b exp(-kt))+e Nelder (1961); Oliver
(1964)
Chapman-Richards f(t) = a(1-b exp(-kt))1/(1-n)
+e Draper dan Smith (1981)
Von Bertalanffy f(t) = (a1-n
-b exp(-kt))1/(1-n)
+e Bertalanffy (1957); Myers
(1986)
Richard’s f(t) = a/(1+b exp(-kt))1/n
+e Richard (1959); Myers
(1986)
Weibull f(t) = (a-b exp(-ktn))+e Ratkowsky (1983); Myers
(1986)
Selain itu, berdasarkan Sitompul dan Guritno (1995), beberapa model untuk
menggambarkan proses pertumbuhan hubungannya dengan umur tanaman adalah
sebagai berikut:
a. Eksponensial tikungan tajam
Pengertian dasar yang perlu dipegang dalam pengembangan model
eksponensial dengan tikungan tajam adalah bahwa proses pertumbuhan itu
disamakan dengan mesin yang dapat menghasilkan suatu produk. Mesin
pertumbuhan itu kemudian dalam tanaman diasumsikan proporsional dengan
biomassa total tanaman. Kemudian mesin tersebut bekerja secara maksimal
sepanjang substrat tersedia, dan pertumbuhan yang dihasilkan tidak dapat balik.
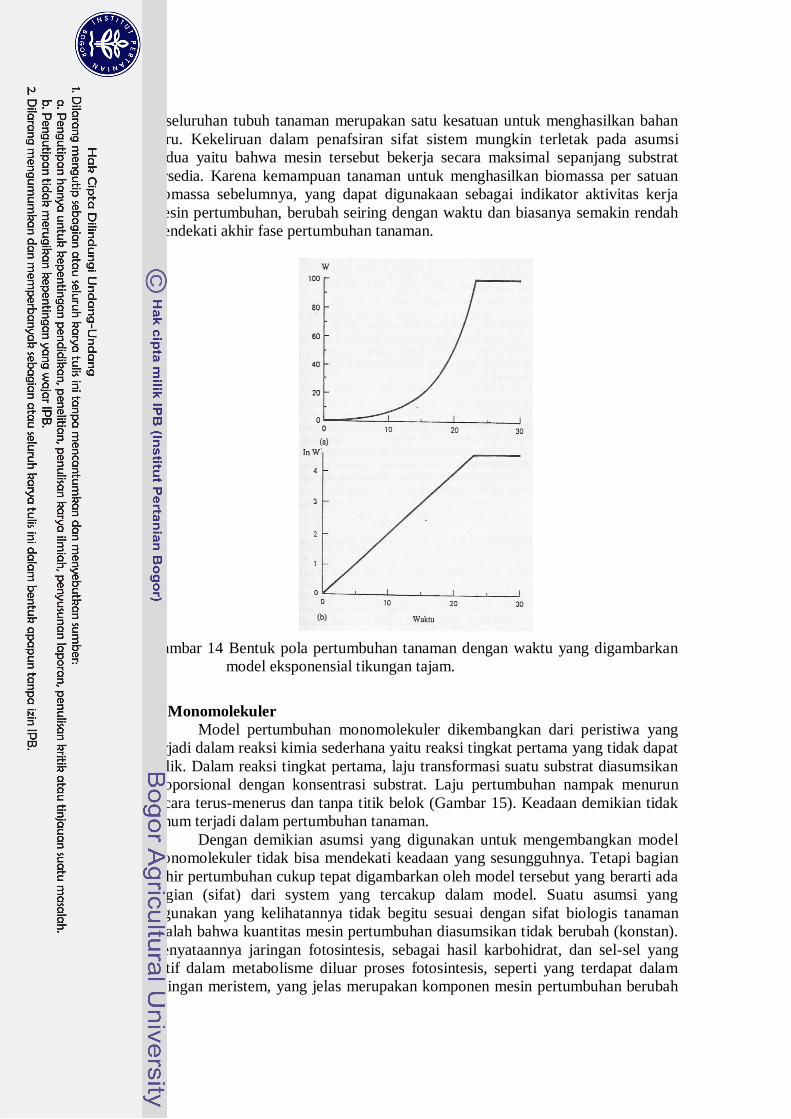
Pertumbuhan dapat berhenti seketika setelah substrat dihabiskan (Gambar 14).
Perkembangan kuantitatif tanaman yang digambarkan model ini sangat
jarang dijumpai khususnya keadaan pertambahan ukuran tanaman yang berhenti
tiba-tiba sebagaimana ditunjukkan oleh tikungan tajam pada model. Memang pada
bagian awal liku, model dapat menstimulasi penampilan tanaman sesungguhnya
yang umumnya mempunyai bentuk pola eksponensial. Ini berarti bahwa asumsi
yang digunakan untuk menurunkan model tersebut hanya dapat mendekati
sebagian proses pertumbuhan sesungguhnya. Asumsi tentang mesin pertumbuhan
yang proporsional dengan biomassa total tanaman cukup realistis, karena
18
keseluruhan tubuh tanaman merupakan satu kesatuan untuk menghasilkan bahan
baru. Kekeliruan dalam penafsiran sifat sistem mungkin terletak pada asumsi
kedua yaitu bahwa mesin tersebut bekerja secara maksimal sepanjang substrat
tersedia. Karena kemampuan tanaman untuk menghasilkan biomassa per satuan
biomassa sebelumnya, yang dapat digunakaan sebagai indikator aktivitas kerja
mesin pertumbuhan, berubah seiring dengan waktu dan biasanya semakin rendah
mendekati akhir fase pertumbuhan tanaman.
Gambar 14 Bentuk pola pertumbuhan tanaman dengan waktu yang digambarkan
model eksponensial tikungan tajam.
b. Monomolekuler
Model pertumbuhan monomolekuler dikembangkan dari peristiwa yang
terjadi dalam reaksi kimia sederhana yaitu reaksi tingkat pertama yang tidak dapat
balik. Dalam reaksi tingkat pertama, laju transformasi suatu substrat diasumsikan
proporsional dengan konsentrasi substrat. Laju pertumbuhan nampak menurun
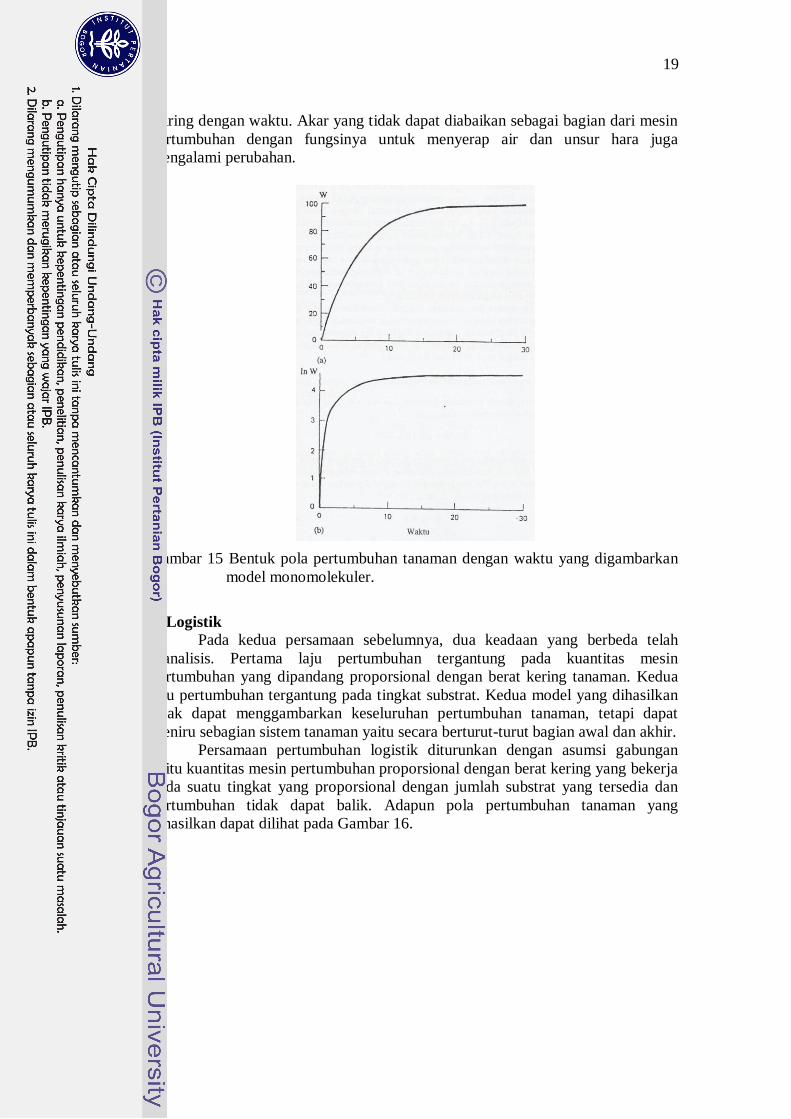
secara terus-menerus dan tanpa titik belok (Gambar 15). Keadaan demikian tidak
umum terjadi dalam pertumbuhan tanaman.
Dengan demikian asumsi yang digunakan untuk mengembangkan model
monomolekuler tidak bisa mendekati keadaan yang sesungguhnya. Tetapi bagian
akhir pertumbuhan cukup tepat digambarkan oleh model tersebut yang berarti ada
bagian (sifat) dari system yang tercakup dalam model. Suatu asumsi yang
digunakan yang kelihatannya tidak begitu sesuai dengan sifat biologis tanaman
adalah bahwa kuantitas mesin pertumbuhan diasumsikan tidak berubah (konstan).
Kenyataannya jaringan fotosintesis, sebagai hasil karbohidrat, dan sel-sel yang
aktif dalam metabolisme diluar proses fotosintesis, seperti yang terdapat dalam
jaringan meristem, yang jelas merupakan komponen mesin pertumbuhan berubah
19
seiring dengan waktu. Akar yang tidak dapat diabaikan sebagai bagian dari mesin
pertumbuhan dengan fungsinya untuk menyerap air dan unsur hara juga
mengalami perubahan.
Gambar 15 Bentuk pola pertumbuhan tanaman dengan waktu yang digambarkan
model monomolekuler.
c. Logistik
Pada kedua persamaan sebelumnya, dua keadaan yang berbeda telah
dianalisis. Pertama laju pertumbuhan tergantung pada kuantitas mesin
pertumbuhan yang dipandang proporsional dengan berat kering tanaman. Kedua
laju pertumbuhan tergantung pada tingkat substrat. Kedua model yang dihasilkan
tidak dapat menggambarkan keseluruhan pertumbuhan tanaman, tetapi dapat
meniru sebagian sistem tanaman yaitu secara berturut-turut bagian awal dan akhir.
Persamaan pertumbuhan logistik diturunkan dengan asumsi gabungan
yaitu kuantitas mesin pertumbuhan proporsional dengan berat kering yang bekerja
pada suatu tingkat yang proporsional dengan jumlah substrat yang tersedia dan
pertumbuhan tidak dapat balik. Adapun pola pertumbuhan tanaman yang
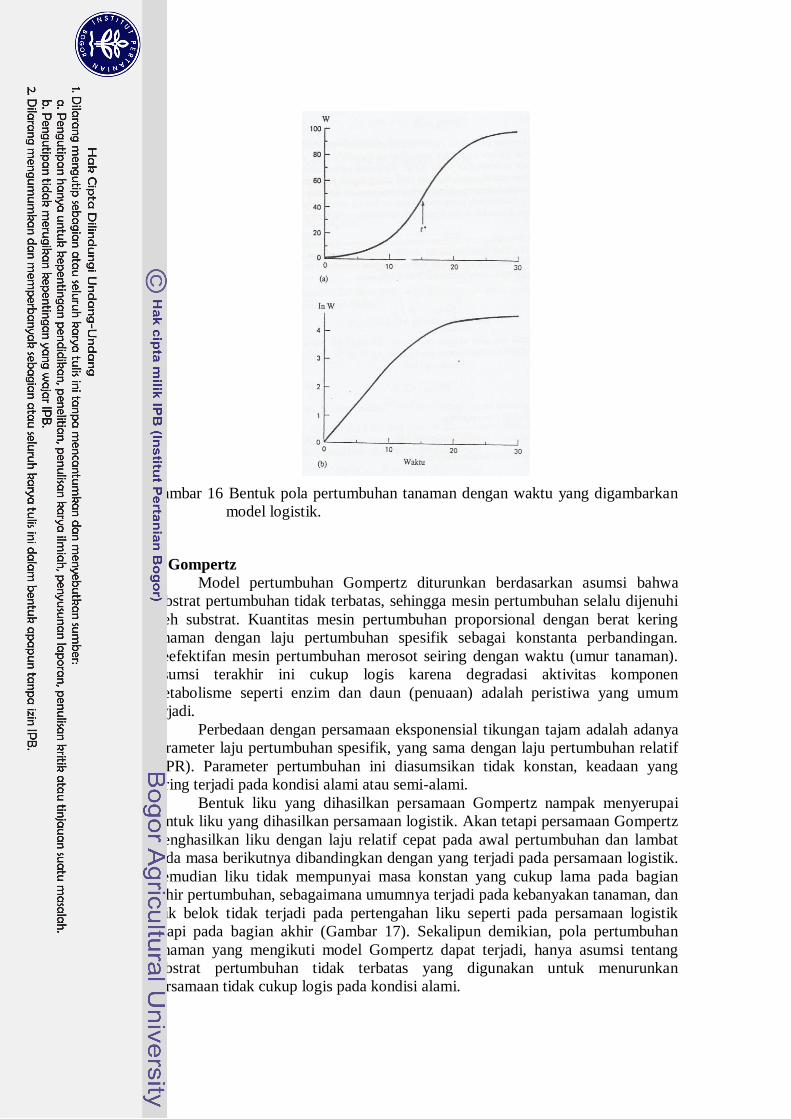
dihasilkan dapat dilihat pada Gambar 16.
20
Gambar 16 Bentuk pola pertumbuhan tanaman dengan waktu yang digambarkan
model logistik.
d. Gompertz
Model pertumbuhan Gompertz diturunkan berdasarkan asumsi bahwa
substrat pertumbuhan tidak terbatas, sehingga mesin pertumbuhan selalu dijenuhi
oleh substrat. Kuantitas mesin pertumbuhan proporsional dengan berat kering
tanaman dengan laju pertumbuhan spesifik sebagai konstanta perbandingan.
Keefektifan mesin pertumbuhan merosot seiring dengan waktu (umur tanaman).
Asumsi terakhir ini cukup logis karena degradasi aktivitas komponen
metabolisme seperti enzim dan daun (penuaan) adalah peristiwa yang umum
terjadi.
Perbedaan dengan persamaan eksponensial tikungan tajam adalah adanya
parameter laju pertumbuhan spesifik, yang sama dengan laju pertumbuhan relatif
(LPR). Parameter pertumbuhan ini diasumsikan tidak konstan, keadaan yang
sering terjadi pada kondisi alami atau semi-alami.
Bentuk liku yang dihasilkan persamaan Gompertz nampak menyerupai
bentuk liku yang dihasilkan persamaan logistik. Akan tetapi persamaan Gompertz
menghasilkan liku dengan laju relatif cepat pada awal pertumbuhan dan lambat
pada masa berikutnya dibandingkan dengan yang terjadi pada persamaan logistik.
Kemudian liku tidak mempunyai masa konstan yang cukup lama pada bagian
akhir pertumbuhan, sebagaimana umumnya terjadi pada kebanyakan tanaman, dan
titik belok tidak terjadi pada pertengahan liku seperti pada persamaan logistik
tetapi pada bagian akhir (Gambar 17). Sekalipun demikian, pola pertumbuhan
tanaman yang mengikuti model Gompertz dapat terjadi, hanya asumsi tentang
substrat pertumbuhan tidak terbatas yang digunakan untuk menurunkan
persamaan tidak cukup logis pada kondisi alami.
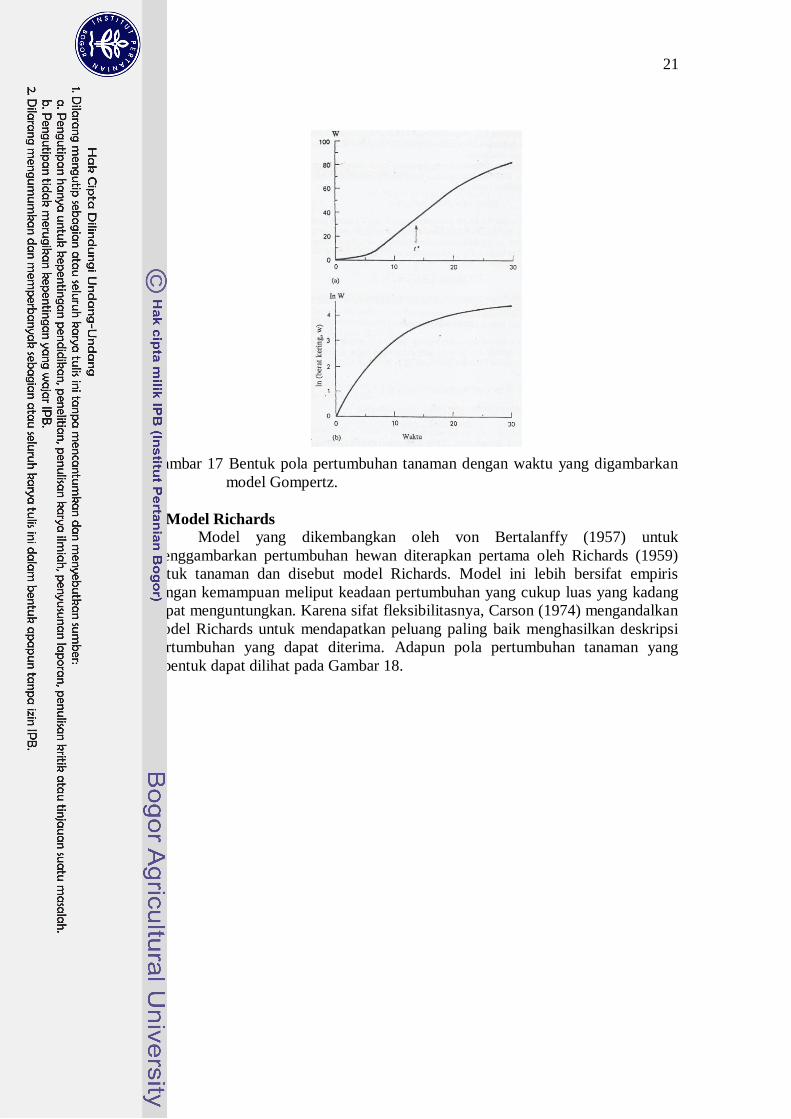
21
Gambar 17 Bentuk pola pertumbuhan tanaman dengan waktu yang digambarkan
model Gompertz.
e. Model Richards
Model yang dikembangkan oleh von Bertalanffy (1957) untuk
menggambarkan pertumbuhan hewan diterapkan pertama oleh Richards (1959)
untuk tanaman dan disebut model Richards. Model ini lebih bersifat empiris
dengan kemampuan meliput keadaan pertumbuhan yang cukup luas yang kadang
dapat menguntungkan. Karena sifat fleksibilitasnya, Carson (1974) mengandalkan
model Richards untuk mendapatkan peluang paling baik menghasilkan deskripsi
pertumbuhan yang dapat diterima. Adapun pola pertumbuhan tanaman yang
dibentuk dapat dilihat pada Gambar 18.
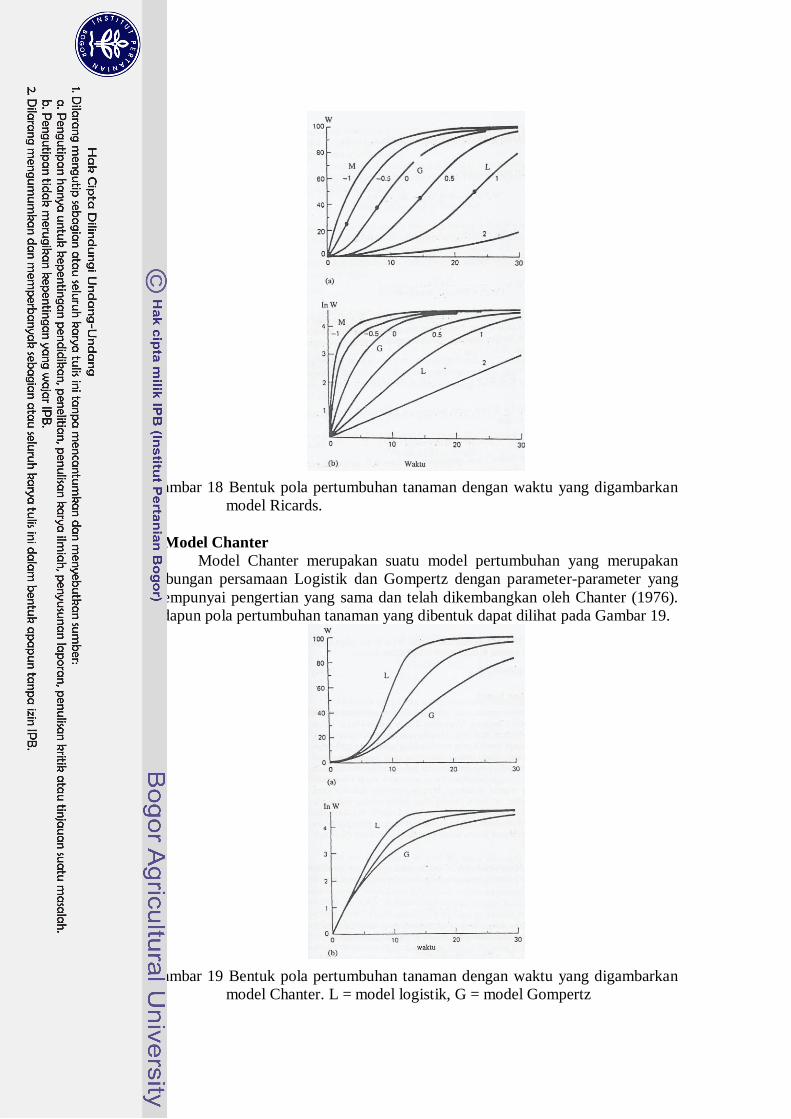
22
Gambar 18 Bentuk pola pertumbuhan tanaman dengan waktu yang digambarkan
model Ricards.
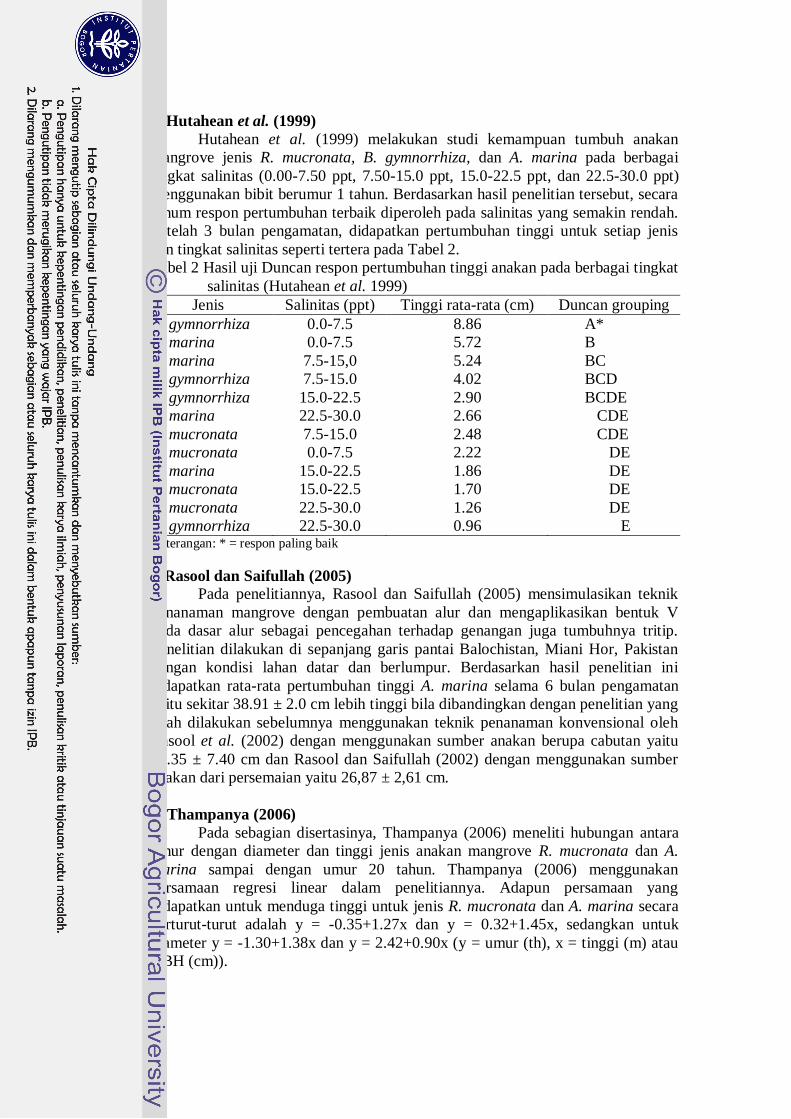
f. Model Chanter
Model Chanter merupakan suatu model pertumbuhan yang merupakan
gabungan persamaan Logistik dan Gompertz dengan parameter-parameter yang
mempunyai pengertian yang sama dan telah dikembangkan oleh Chanter (1976).
Adapun pola pertumbuhan tanaman yang dibentuk dapat dilihat pada Gambar 19.
Gambar 19 Bentuk pola pertumbuhan tanaman dengan waktu yang digambarkan
model Chanter. L = model logistik, G = model Gompertz

23
Penelitian Pertumbuhan Mangrove
Terdapat beberapa penelitian yang mengkaji mengenai pertumbuhan
mangrove khusunya untuk jenis A. marina dan R. mucronata diantaranya sebagai
berikut:
a. Burchett et al. (1984)
Burchett et al. (1984) meneliti hubungan antara parameter pertumbuhan
dan respirasi akar A. marina dengan berbagai tingkat salinitas (0%, 25%, 75%,
dan 100% air laut). Pertumbuhan (biomassa dan luas permukaan daun) dan rata-
rata respirasi tertinggi didapatkan pada media 25% air laut, tingkat sukulensi daun
tertinggi pada media 50% air laut, dan potensial osmotik daun tertinggi pada
media 100% air laut.
b. O’Grady et al. (1996)
O’Grady et al. (1996) meneliti pertumbuhan dan distribusi dari dua jenis
anakan mangrove (A. marina dan R. stylosa) di area pantai Darwin Harbour.
Berdasarkan penelitian tersebut, anakan A. marina dan R. stylosa memiliki tingkat
kerapatan dan pertumbuhan terbesar pada areal dengan kanopi yang terbuka.
Secara umum anakan R. stylosa lebih tahan bila dibandingkan dengan A. marina.
Rhizophora memiliki cadangan embrionik yang lebih besar dibandingkan
Avicennia. Hal ini memungkinkan anakan Rhizophora dapat lebih bertahan di
bawah naungan untuk periode yang lama dibandingkan dengan Avicennia.
c. Devoe dan Cole (1998)
Devoe dan Cole (1998) melakukan penelitian mengenai pertumbuhan
hutan mangrove di Federated States of Micronesia (FSM). Berdasarkan plot
permanen yang telah dibangun selama 9 tahun di lokasi ini, didapatkan nilai riap
rat-rata tahunan dari jenis R. apiculata sekitar 0.25 cm/th, Xylocarpus granatum
sekitar 0.31 cm/th, R. mucronata sekitar 0.37 cm/th, B. gymnorrhiza sekitar 0.35
cm/th, dan S. alba sekitar 0.49 cm/th. Secara keseluruhan, riap volume rata-rata di
area FSM ini mencapai 4.5 m3/(ha th).
d. Komiyama et al. (1998)
Komiyama et al. (1998) mengujicobakan penanaman R. apiculata dan R.
mucronata dengan teknik stek propagul. Propagul masing-masing jenis dibagi
menjadi tiga bagian yaitu bawah, tengah, dan atas. Berdasarkan penelitian
tersebut, rata-rata tinggi batang dan diameter untuk jenis R. mucronata terbesar
dihasilkan oleh potongan propagul bagian bawah, kemudian sedang untuk
propagul bagian tengah, dan terkecil untuk propagul bagian atas. Pada jenis R.
apiculata, rata-rata diameter yang dihasilkan memiliki kecenderungan yang sama
dengan jenis R. mucronata, akan tetapi rata-rata tinggi batang tidak berbeda untuk
ketiga bagian propagul yang digunakan. Setelah 38 bulan penanaman, rata-rata
diameter untuk jenis R. apiculata dan R. mucronata secara berturut-turut 0.78-
1.37 dan 0.56-0.89 kali rata-rata diameter anakan yang berasal dari propagul utuh,
sedangkan untuk rata-rata tinggi 0.81-0.85 dan 0.50-1.00 kali.
24
e. Hutahean et al. (1999)
Hutahean et al. (1999) melakukan studi kemampuan tumbuh anakan
mangrove jenis R. mucronata, B. gymnorrhiza, dan A. marina pada berbagai
tingkat salinitas (0.00-7.50 ppt, 7.50-15.0 ppt, 15.0-22.5 ppt, dan 22.5-30.0 ppt)
menggunakan bibit berumur 1 tahun. Berdasarkan hasil penelitian tersebut, secara
umum respon pertumbuhan terbaik diperoleh pada salinitas yang semakin rendah.
Setelah 3 bulan pengamatan, didapatkan pertumbuhan tinggi untuk setiap jenis
dan tingkat salinitas seperti tertera pada Tabel 2.
Tabel 2 Hasil uji Duncan respon pertumbuhan tinggi anakan pada berbagai tingkat
salinitas (Hutahean et al. 1999)
Jenis Salinitas (ppt) Tinggi rata-rata (cm) Duncan grouping
B. gymnorrhiza 0.0-7.5 8.86 A*
A. marina 0.0-7.5 5.72 B
A. marina 7.5-15,0 5.24 BC
B. gymnorrhiza 7.5-15.0 4.02 BCD
B. gymnorrhiza 15.0-22.5 2.90 BCDE
A. marina 22.5-30.0 2.66 CDE
R. mucronata 7.5-15.0 2.48 CDE
R. mucronata 0.0-7.5 2.22 DE
A. marina 15.0-22.5 1.86 DE
R. mucronata 15.0-22.5 1.70 DE
R. mucronata 22.5-30.0 1.26 DE
B. gymnorrhiza 22.5-30.0 0.96 E Keterangan: * = respon paling baik
f. Rasool dan Saifullah (2005)
Pada penelitiannya, Rasool dan Saifullah (2005) mensimulasikan teknik
penanaman mangrove dengan pembuatan alur dan mengaplikasikan bentuk V
pada dasar alur sebagai pencegahan terhadap genangan juga tumbuhnya tritip.
Penelitian dilakukan di sepanjang garis pantai Balochistan, Miani Hor, Pakistan
dengan kondisi lahan datar dan berlumpur. Berdasarkan hasil penelitian ini
didapatkan rata-rata pertumbuhan tinggi A. marina selama 6 bulan pengamatan
yaitu sekitar 38.91 ± 2.0 cm lebih tinggi bila dibandingkan dengan penelitian yang
telah dilakukan sebelumnya menggunakan teknik penanaman konvensional oleh
Rasool et al. (2002) dengan menggunakan sumber anakan berupa cabutan yaitu
12.35 ± 7.40 cm dan Rasool dan Saifullah (2002) dengan menggunakan sumber
anakan dari persemaian yaitu 26,87 ± 2,61 cm.
g. Thampanya (2006)
Pada sebagian disertasinya, Thampanya (2006) meneliti hubungan antara
umur dengan diameter dan tinggi jenis anakan mangrove R. mucronata dan A.
marina sampai dengan umur 20 tahun. Thampanya (2006) menggunakan
persamaan regresi linear dalam penelitiannya. Adapun persamaan yang
didapatkan untuk menduga tinggi untuk jenis R. mucronata dan A. marina secara
berturut-turut adalah y = -0.35+1.27x dan y = 0.32+1.45x, sedangkan untuk
diameter y = -1.30+1.38x dan y = 2.42+0.90x (y = umur (th), x = tinggi (m) atau
DBH (cm)).

25
h. Jumiati (2008)
Jumiati (2008) melakukan penelitian mengenai pertumbuhan R. mucronata
dan R. apiculata di kawasan yang terpolusi oleh minyak di kawasan tambang
minyak dan gas PT Medco E & P di kecamatan Tarakan Timur. Pengukuran
dilakukan dengan interval 2 minggu selama 4 bulan pengamatan pada tiga zona
berbeda yaitu zona darat, tengah, dan laut. Anakan yang digunakan dalam
penelitian ini berupa propagul dan bibit. Adapun pertambahan tinggi rata-rata
semai R. mucronata dan R. apiculata yang didapatkan tertera pada Tabel 3.
Tabel 3 Pertambahan tinggi rata-rata semai (cm) R. mucronata (Rm) dan R.
apiculata (Ra) pada zona darat, tengah, dan laut di tapak Medco E&P Parameter Riap tinggi rata-rata (cm/2 minggu)
R. mucronata R. apiculata
Bibit Propagul Bibit Propagul
D T L D T L D T L D T L
Pertambahan
tinggi (cm) 5.2 1.6 7.2 35.7 6.1 18.5 1.9 0.34 4.9 8.7 7.6 15.7
Keterangan: D = zona darat, T = zona tengah, L = zona laut
i. Kairo et al. (2008)
Kairo et al. (2008) melakukan penelitian mengenai struktur dan
produktivitas dari hutan tanaman R. mucronata berumur 12 tahun di Gazi Bay,
Kenya. Berdasarkan hasil penelitiannya didapatkan bahwa rata-rata tinggi kanopi
dari R. mucronata berumur 12 tahun yaitu 8.4 ± 1.1 m dengan rata-rata diameter
6.2 ± 1.87 cm. Biomassanya diperkirakan mencapai 106.7 ± 24.0 ton/ha dengan
akumulasi biomassa rata-rata 8.9 ton/(ha th).
k. Halidah (2010)
Halidah (2010) meneliti tentang pengaruh tinggi genangan dan jarak tanam
terhadap pertumbuhan anakan R. mucronata di pantai barat Sulawesi Selatan.
Berdasarkan penelitian tersebut, perlakuan tinggi genangan belum menunjukkan
pengaruh terhadap pertumbuhan tinggi, sedangkan jarak tanam memberikan
pengaruh yang sangat nyata terhadap pertumbuhan tinggi. Jarak tanam 0.5 m x 0.5
m, 1 m x 1 m, 1 m x 2 m, dan 2 m x 1.5 m memberikan rata-rata tinggi R.
mucronata berumur 6 bulan secara berturut-turut yaitu 1.56 cm, 2.22 cm, 1.77 cm,
dan 5.74 cm.
l. Syah (2011)
Syah (2011) melakukan penelitian mengenai pertumbuhan tanaman bakau
(R. mucronata) pada lahan restorasi di hutan lindung Angke Kapuk provinsi DKI
Jakarta. Berdasarkan penelitian tersebut, pada umur 3 bulan, rata-rata tinggi R.
mucronata berkisar antara 60.05-60.39 cm, sedangkan rata-rata diameter berkisar
antara 3.33-3.90 cm. Pada umur 6 bulan, rata-rata tinggi berkisar antara 104.90-
106.00 cm, sedangkan rata-rata diameter berkisar antara 3.33-3.90 cm. Pada umur
12 bulan, rata-rata tinggi dan diameter berturut-turut berkisar antara 140.65-
142.82 cm dan 5.63-5.75 cm. Sedangkan pada umur 16 bulan, rata-rata tinggi dan
diameter berturut-turut berkisar antara 129.48-148.82 cm dan 6.08-6.19 cm.