







Bahasa
Halaman
Undang-undang


Annals of West University of Timişoara, ser. Biology, 2014, vol XVII (2), pp.111-122
111
PROPERTIES OF AMINOTRANSFERASES FROM
TELADORSAGIA CIRCUMCINCTA
Noorzaid MUHAMAD
a, David C. SIMCOCK
bcd, Heather V. SIMPSON
e,
Kevin C. PEDLEYb, Simon BROWN
cf
aUniversity Kuala Lumpur, Royal College of Medicine Perak, Ipoh, Perak, Malaysia, bInstitute of Food, Nutrition and Human Health, Massey University, New Zealand,
cDeviot Institute, Deviot, Tasmania 7275, Australia,
dFaculty of Medicine, Health and Molecular Sciences, James Cook University, Australia,
eInstitute of Veterinary and Biomedical Sciences, Massey University, New Zealand,
fSchool of Human Life Sciences, University of Tasmania, Launceston, Tasmania 7250,
Australia
Corresponding author e-mail: [email protected] Received 30 August 2014; accepted 4 November 2014
ABSTRACT Activities of several aminotransferases were measured in L3 and adult Teladorsagia
circumcincta, but most of these had maximal activity of less than 8 nmol min-1
mg-1
protein. Only aspartate aminotransferase (AspAT) and alanine aminotransferase
(AlaAT) activities exceeded this value and some kinetic properties of these enzymes
were characterised. For L3 AspAT, the apparent Kms were 1.2 mM, 0.13 mM, 0.11 mM
and 0.04 mM for aspartate, α-ketoglutarate, glutamate and oxaloacetate, respectively,
and the apparent Vmaxs were 960 nmol min-1
mg-1
protein for aspartate deamination
and 420 nmol min-1
mg-1
protein in the direction of glutamate deamination. For L3
AlaAT, the apparent Kms were 5.2 mM, 0.5 mM, 0.5 mM and 1.2 mM for alanine, α-
ketoglutarate, glutamate and pyruvate, respectively, and apparent Vmaxs were 107
nmol min-1
mg-1
protein for alanine deamination and 48 nmol min-1
mg-1
protein for
alanine formation. Both enzymes required exogenous pyridoxal 5′-phosphate for
optimal activity. The equilibrium constants for the AspAT and AlaAT reactions were
consistent with those estimated from the estimated kinetic parameters. From these
parameters we infer that T. circumcincta AlaAT is present predominantly as a
mitochondrial enzyme favouring pyruvate formation while AspAT is predominantly a
cytosolic enzyme favouring glutamate formation.
KEY WORDS: aminotransferase, enzyme kinetics, nematode, parasite.
INTRODUCTION
Teladorsagia circumcincta is a nematode parasite of sheep that secretes or
excretes NH3/NH4+ (Muhamad et al., 2012) and urea (Muhamad et al., 2013), but
appears not to secrete or excrete amino acids (Muhamad, 2006), although secretion or
excretion of all three has been reported for other helminths (Barrett, 1991). We have
characterised the kinetics of some of the enzymes that might be involved in generating
these compounds (Muhamad et al., 2005; Muhamad et al., 2011). However, it is also

MUHAMAD et al: Some properties of aminotransferases from Teladorsagia circumcincta
112
important to consider the intracellular transfer of nitrogenous groups that provides
alternative connections between the metabolites involved in these reactions (Muhamad
et al., 2005; Muhamad et al., 2011). Arguably, transaminations are among the most
important of these and, of those, the reactions catalysed by aspartate aminotransferase
(AspAT, E.C. 2.6.1.1)
aspartate + α-ketoglutarate ⇌ glutamate + oxaloacetate
and alanine aminotransferase (AlaAT, E.C. 2.6.1.2)
alanine + α-ketoglutarate ⇌ glutamate + pyruvate
are of particular significance, although other aminotransferases have been detected in
nematodes (Barrett, 1991).
Both AspAT and AlaAT catalyse the transfer of an amino group from an
amino acid donor (aspartate or alanine, respectively) to a ketoacid acceptor (α-
ketoglutarate) generating an amino acid (glutamate) and a different ketoacid
(oxaloacetate or pyruvate, respectively). The reactions are readily reversible, as
is apparent from ∆rG´0 = 4.96 kJ mol
-1 for AspAT (Kishore et al., 1998) and
∆rG´0 = 0.46 kJ mol
-1 for AlaAT (Tewari et al., 1998). Both AspAT and
AlaAT require pyridoxal 5′-phosphate (PLP), which can be lost from the
enzyme and incubation of the enzyme suspension with PLP increases the
observed rate of reaction as the PLP is reincorporated into the apoenzyme
(Martinez-Carrion et al., 1967; Scardi et al., 1963). Aminotransferases, including AlaAT and AspAT, have been detected in
several helminth species (Barrett, 1991), but the only data for T. circumcincta appears
to be the estimate of AlaAT activity given by Walker and Barrett (1991). Most of the
reports for other species also provide only an estimate of the activity. For example,
Grantham and Barrett (1986) surveyed the activity of many aminotransferases in
Heligmosomoides polygyrus and Panagrellus redivivus without providing any other
indication of the kinetics of each reaction. This is not enough to begin to understand
the interactions between the reactions producing nitrogenous compounds (Muhamad et
al., 2005; Muhamad et al., 2011). In a small number of cases more data are available.
For example, Sharma and Singh (1977) reported apparent Kms for aspartate of 9 mM
and 2.9 mM aspartate for AspAT from male and female, respectively, Stephanurus
dentatus and activity was greatest at pH 8 for both genders. Burenina (2001) reported
the apparent Kms for alanine and α-ketoglutarate of both the mitochondrial and
cytoplasmic isozymes of AlaAT from the trematode Eurytrema pancreaticum. Lowe
and Rowe (1985) reported the Kms for aspartate and α-ketoglutarate of the single form
of AspAT they could identify in the protozoon Trichomonas vaginalis, later they
reported a second isoform that differed only in the Vmax for substrates other than

Annals of West University of Timişoara, ser. Biology, 2014, vol XVII (2), pp.111-122
113
aspartate (Lowe & Rowe, 1986). Here we describe some of the kinetic properties of
AspAT and AlaAT from T. circumcincta.
MATERIAL AND METHOD
Parasite culture and homogenate preparation
Pure strains of Teladorsagia circumcincta were maintained by regular passage
through sheep to provide L3. Larvae were concentrated by centrifugation at
approximately 1000 × g, washed and then resuspended in 100 mM KH2PO4-KOH pH
7.5. Where necessary adult T. circumcincta were obtained from the abomasal contents
of donor sheep as described previously (Simcock et al., 2006). It was not practicable to
culture sufficient L3 or obtain enough adults to purify the enzymes and so all the work
described here was carried out using nematode homogenates. In order to homogenise
the nematodes, the suspension was frozen at -20°C and then ground manually at 4°C as
described previously (Walker et al., 2007).
Enzyme activities
The activity of AspAT and AlaAT were assayed by coupling the reaction with
an appropriate NAD+-linked dehydrogenase. Each reaction was monitored
spectrophotometrically by following the oxidation of NADH at 340 nm (ε340 = 6220
M−1
cm−1
) using 50 µg mL−1
protein at 30°C. All assays were carried out using either
an Ultrospec III (LKB Instruments) or a CE 599 (Cecil Instruments Ltd., Cambridge,
England) spectrophotometer from which the data were collected by computer using
software and hardware developed in-house as previously reported (Walker et al.,
2007). For enzymes other than AspAT and AlaAT, for which the aminotransferse
activities were very low, the rates were estimated discontinuously over 30-60 min. In
all cases data from at least three homogenates were analysed (n ≥ 3), except for the pH
dependence experiments where the data represent only two separate homogenates (n =
2).
The activity of AspAT was determined in the direction of aspartate
deamination by coupling the reaction to malate dehydrogenase (MDH, E. C. 1.1.1.37)
so that the oxaloacetate produced was reduced to malate and the concentration of
NADH declined. Unless otherwise noted, the assay mixture consisted of 10 mM
aspartate and 0.2 mM α-ketoglutarate, 200 µM NADH, 200 µM PLP, 1 U mL-1
MDH,
50 µg mL-1
homogenate protein, 100 mM Tris-HCl pH 7.5. In the direction of aspartate
formation by AspAT, the α-ketoglutarate generated was reductively aminated to
glutamate by glutamate dehydrogenase (GDH, E. C. 1.4.1.3) and the concentration of
NADH declined. Unless otherwise specified, the assay mixture consisted of 1 mM
glutamate, 2 mM oxaloacetate, 200 µM NADH, 200 µM PLP, 1 U mL-1
GDH, 50 µg
mL-1
homogenate protein, 100 mM Tris-HCl pH 7.5. Teladorsagia circumcincta has
considerable endogenous activities of both MDH (Simcock et al., 2012) and GDH

MUHAMAD et al: Some properties of aminotransferases from Teladorsagia circumcincta
114
(Muhamad et al., 2011) in addition to the exogenous enzyme provided. Further details
of the assays are provided in the figure legends.
The activity of AlaAT was determined in the direction of alanine deamination
by coupling the reaction to lactate dehydrogenase (LDH, E. C. 1.1.1.27) so that the
pyruvate produced was reduced to lactate and the decline in the concentration of
NADH was monitored at 340 nm. Unless otherwise noted, the assay mixture consisted
of 10 mM alanine and 2 mM α-ketoglutarate and 200 µM NADH, 200 µM PLP, 1 U
mL-1
LDH, 50 µg mL-1
homogenate protein, 100 mM Tris-HCl pH 7.5. In the direction
of alanine formation by AlaAT, the α-ketoglutarate generated was reductively
aminated to glutamate by GDH and the reaction was monitored continuously by
following the decline in the the absorbance at 340 nm as a measure of the
concentration of NADH. Unless otherwise specified, the assay mixture consisted of 5
mM glutamate and 2 mM pyruvate and 200 µM NADH, 200 µM PLP, 40 mM NH4Cl,
1 U mL-1
GDH, 50 µg mL-1
homogenate protein, 100 mM Tris-HCl pH 7.5.
Teladorsagia circumcincta has considerable endogenous activities of both MDH
(Simcock et al., 2012) and GDH (Muhamad et al., 2011) in addition to the exogenous
enzyme provided. Further details of the assays are provided in the figure legends.
The protein concentration of homogenates was determined using the Bradford
method (Bradford, 1976) and bovine serum albumin as the standard. No detergents
were employed to solubilise the nematode homogenate as they can interfere with
protein assays (Sapan et al., 1999).
Data analysis
While it is likely that the kinetics of L3 T. circumcincta AspAT and AlaAT can
be modelled using the same mechanisms as those of other species (Cascante & Cortés,
1988; Creighton & Murthy, 1990), we have not attempted to employ such models in
the analysis presented here. Instead, apparent Kms and Vmaxs were obtained by fitting
the standard Michaelis-Menten expression to the activity data for each substrate. This
approach is consistent with that adopted previously (Muhamad et al., 2011). Estimates
of apparent Vmax depend on the enzyme concentration, so comparisons between L3 and
adult should be made with caution, but such comparisons can be made more reliably
for estimates involving the same lifecycle stage. Estimates of apparent Km are
independent of enzyme concentration, so comparisons between values for L3 and adult
preparations can be made with more confidence. All nonlinear regression and the other
analyses described below were carried out using R (Ihaka & Gentleman, 1996).
RESULTS
In the direction of aspartate deamination the apparent Kms for aspartate and α-
ketoglutarate of L3 AspAT were 1.2 ± 0.2 mM and 0.13 ± 0.02 mM, respectively, and
the apparent Vmax was 960 ± 50 nmol min-1
mg-1
protein (Figure 1, A and B). In the
direction of glutamate deamination the apparent Kms for glutamate and oxaloacetate
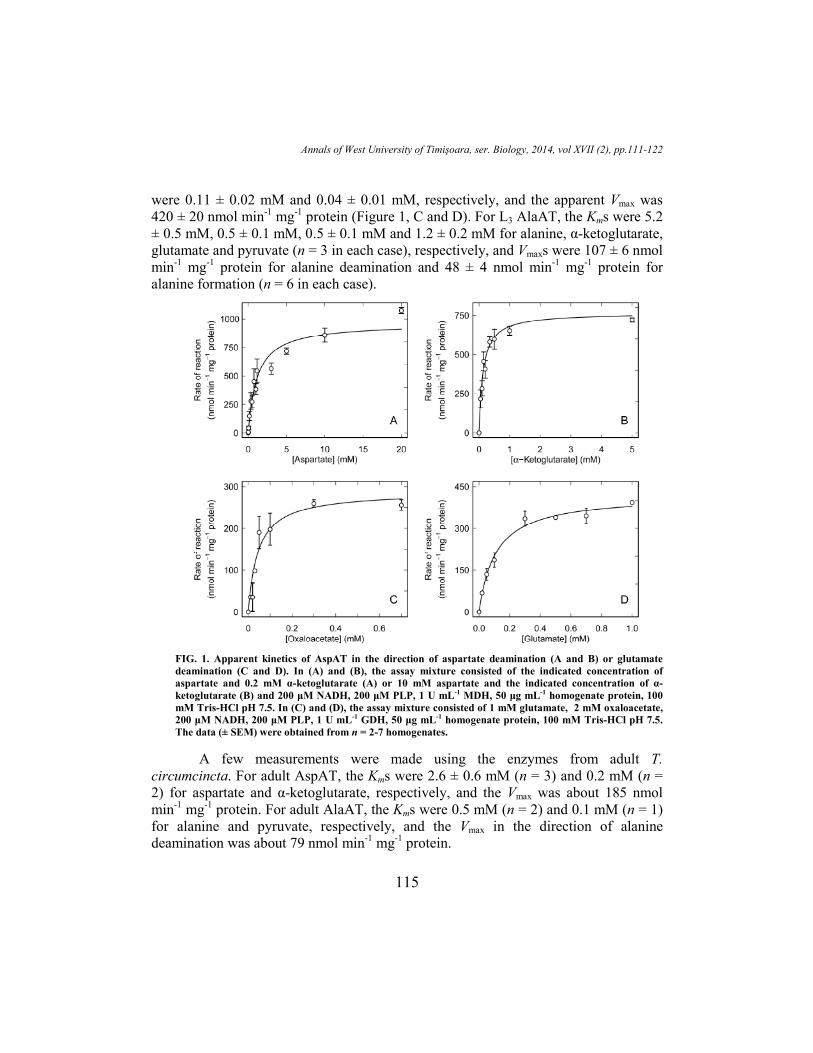
Annals of West University of Timişoara, ser. Biology, 2014, vol XVII (2), pp.111-122
115
were 0.11 ± 0.02 mM and 0.04 ± 0.01 mM, respectively, and the apparent Vmax was
420 ± 20 nmol min-1
mg-1
protein (Figure 1, C and D). For L3 AlaAT, the Kms were 5.2
± 0.5 mM, 0.5 ± 0.1 mM, 0.5 ± 0.1 mM and 1.2 ± 0.2 mM for alanine, α-ketoglutarate,
glutamate and pyruvate (n = 3 in each case), respectively, and Vmaxs were 107 ± 6 nmol
min-1
mg-1
protein for alanine deamination and 48 ± 4 nmol min-1
mg-1
protein for
alanine formation (n = 6 in each case).
FIG. 1. Apparent kinetics of AspAT in the direction of aspartate deamination (A and B) or glutamate
deamination (C and D). In (A) and (B), the assay mixture consisted of the indicated concentration of
aspartate and 0.2 mM α-ketoglutarate (A) or 10 mM aspartate and the indicated concentration of α-
ketoglutarate (B) and 200 µM NADH, 200 µM PLP, 1 U mL-1 MDH, 50 µg mL-1 homogenate protein, 100
mM Tris-HCl pH 7.5. In (C) and (D), the assay mixture consisted of 1 mM glutamate, 2 mM oxaloacetate,
200 µM NADH, 200 µM PLP, 1 U mL-1 GDH, 50 µg mL-1 homogenate protein, 100 mM Tris-HCl pH 7.5.
The data (± SEM) were obtained from n = 2-7 homogenates.
A few measurements were made using the enzymes from adult T.
circumcincta. For adult AspAT, the Kms were 2.6 ± 0.6 mM (n = 3) and 0.2 mM (n =
2) for aspartate and α-ketoglutarate, respectively, and the Vmax was about 185 nmol
min-1
mg-1
protein. For adult AlaAT, the Kms were 0.5 mM (n = 2) and 0.1 mM (n = 1)
for alanine and pyruvate, respectively, and the Vmax in the direction of alanine
deamination was about 79 nmol min-1
mg-1
protein.

MUHAMAD et al: Some properties of aminotransferases from Teladorsagia circumcincta
116
The PLP prosthetic group of an aminotransferase is often labile, but it can be
reincorporated when added to an enzyme suspension (Martinez-Carrion et al., 1967).
In the case of AspAT, the rate of glutamate deamination was increased to 375 nmol
min-1
mg-1
protein after incubation with 0.1 mM PLP (Figure 2A). The activity of
AspAT was almost completely inhibited by 1 mM PLP. The activity of L3 AlaAT was
more than doubled by incubation with PLP (Figure 2B) and the enzyme was more
sensistive than AspAT to concentrations of PLP in the range 0.2 mM to 0.5 mM
(Figure 2, A and B).
FIG. 2. The effect of added pyridoxal 5′-phosphate (PLP) on the rate of aspartate (A) or alanine (B)
deamination. The assays were carried out using 3 mM aspartate, 1 mM α-ketoglutarate, 200 µM NADH, 1
U mL-1 MDH, 50 µg mL-1 homogenate protein, 100 mM Tris-HCl pH 7.5. The homogenate was incubated
with the indicated concentration of PLP in the reaction mixture for 2 minutes before the reaction was
initiated by the addition of α-ketoglutarate.
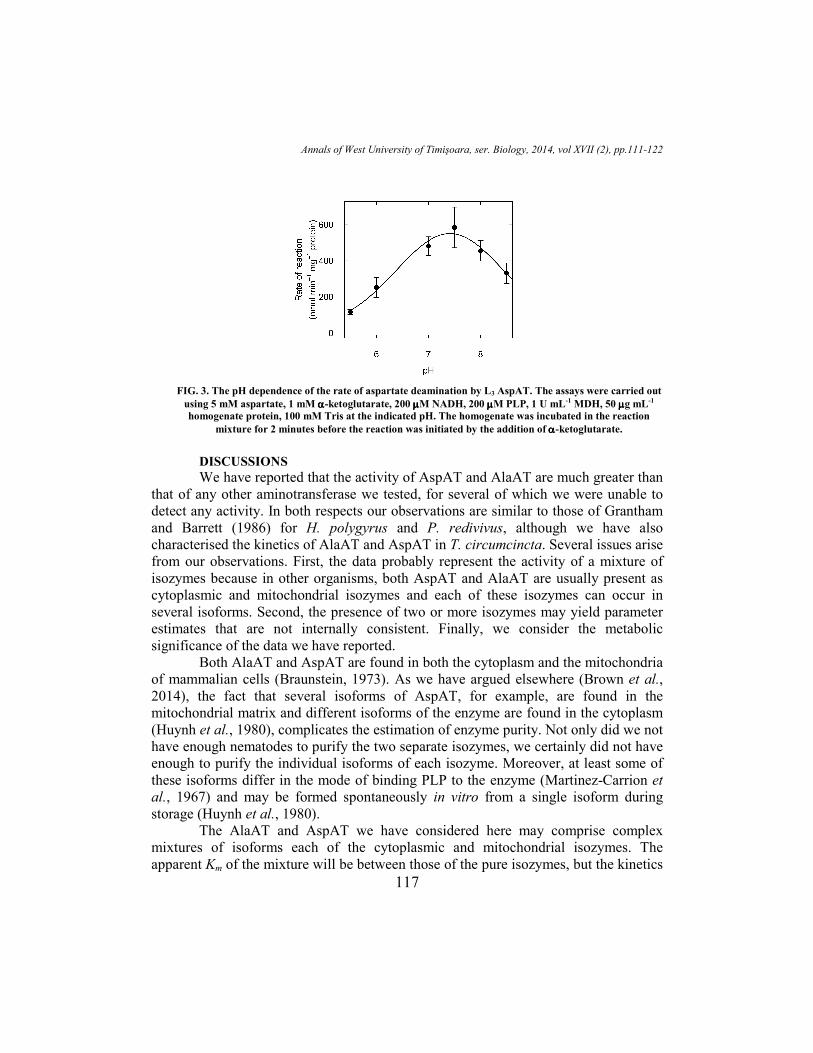
The aspartate deamination activity of L3 AspAT was greatest at pH 7.4 and
declined at lower pH with an apparent pKa of 6.2 and at higher pH with an apparent
pKa of 8.7 (Figure 3). The low activity of L3 AlaAT made it relatively difficult to
assess the pH sensitivity of the enzyme, but the rate of alanine deamination appeared to
be greatest at about pH 7.5.
We also attempted to measure the activity of several other aminotransferases
using α-ketoglutarate as the ketoacid acceptor, but in L3 T. circumcincta the rates were
less than about 2 nmol min-1
mg-1
protein using valine, histidine, leucine or glycine,
and no activity could be detected using cysteine, isoleucine or ornithine (the lower
limit of detection was about 0.1 nmol min-1
mg-1
protein). Greater activity (up to about
8 nmol min-1
mg-1
protein) was observed for serine and valine using pyruvate as the
ketoacid acceptor. In adult nematode homogenates rates of less than about 2 nmol min-
1 mg
-1 protein were observed using cysteine, valine, isoleucine, histidine and lysine
using α-ketoglutarate as the ketoacid acceptor.
Annals of West University of Timişoara, ser. Biology, 2014, vol XVII (2), pp.111-122
117
FIG. 3. The pH dependence of the rate of aspartate deamination by L3 AspAT. The assays were carried out
using 5 mM aspartate, 1 mM αααα-ketoglutarate, 200 µµµµM NADH, 200 µµµµM PLP, 1 U mL-1 MDH, 50 µµµµg mL-1
homogenate protein, 100 mM Tris at the indicated pH. The homogenate was incubated in the reaction
mixture for 2 minutes before the reaction was initiated by the addition of αααα-ketoglutarate. DISCUSSIONS
We have reported that the activity of AspAT and AlaAT are much greater than
that of any other aminotransferase we tested, for several of which we were unable to
detect any activity. In both respects our observations are similar to those of Grantham
and Barrett (1986) for H. polygyrus and P. redivivus, although we have also
characterised the kinetics of AlaAT and AspAT in T. circumcincta. Several issues arise
from our observations. First, the data probably represent the activity of a mixture of
isozymes because in other organisms, both AspAT and AlaAT are usually present as
cytoplasmic and mitochondrial isozymes and each of these isozymes can occur in
several isoforms. Second, the presence of two or more isozymes may yield parameter
estimates that are not internally consistent. Finally, we consider the metabolic
significance of the data we have reported.
Both AlaAT and AspAT are found in both the cytoplasm and the mitochondria
of mammalian cells (Braunstein, 1973). As we have argued elsewhere (Brown et al.,
2014), the fact that several isoforms of AspAT, for example, are found in the
mitochondrial matrix and different isoforms of the enzyme are found in the cytoplasm
(Huynh et al., 1980), complicates the estimation of enzyme purity. Not only did we not
have enough nematodes to purify the two separate isozymes, we certainly did not have
enough to purify the individual isoforms of each isozyme. Moreover, at least some of
these isoforms differ in the mode of binding PLP to the enzyme (Martinez-Carrion et
al., 1967) and may be formed spontaneously in vitro from a single isoform during
storage (Huynh et al., 1980).
The AlaAT and AspAT we have considered here may comprise complex
mixtures of isoforms each of the cytoplasmic and mitochondrial isozymes. The
apparent Km of the mixture will be between those of the pure isozymes, but the kinetics

MUHAMAD et al: Some properties of aminotransferases from Teladorsagia circumcincta
118
of such mixtures tend to resemble those of a homogeneous population of enzymes
unless the Kms of the isoforms are sufficiently different (Brown et al., 2014). For
example, we have estimated that mitcyt
mmKK=β (or β
-1) must be at least 20 for the
isoforms to be apparent from the kinetics of the mixture (Brown et al., 2014). This is
rarely the case for AspAT (the pig heart enzyme is one example where β > 20
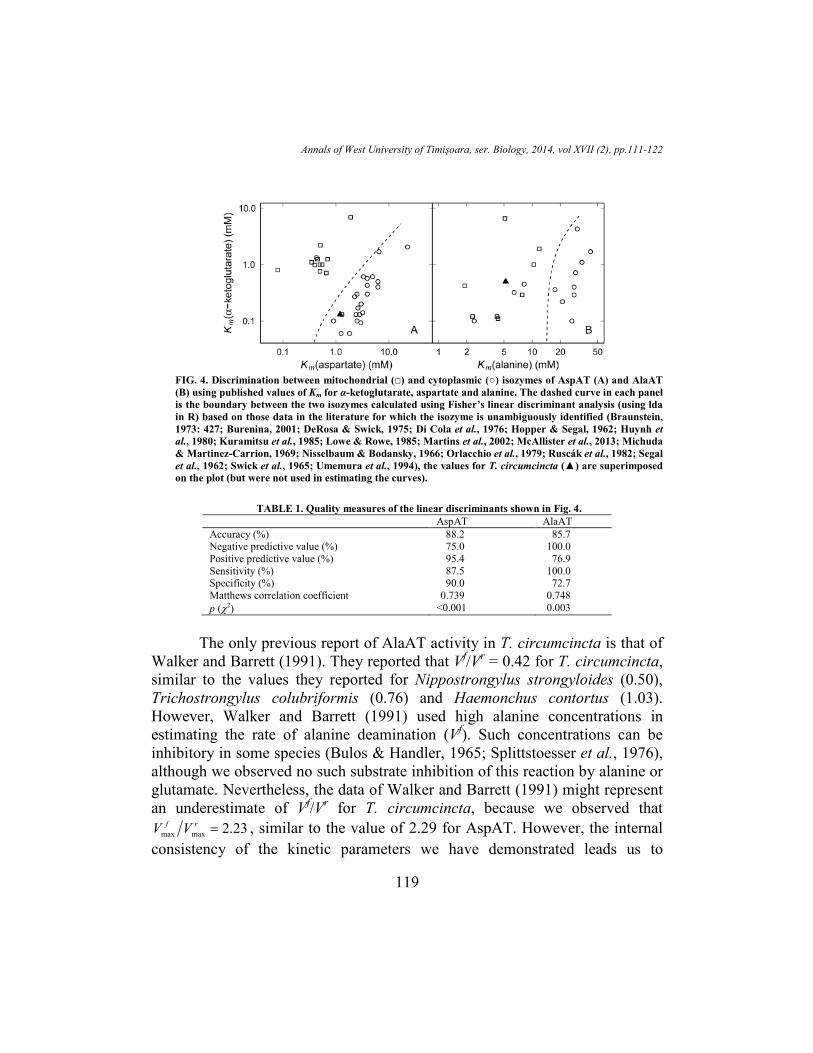
(Braunstein, 1973: 427), although some preparations of these isozymes give β < 20
(Nisselbaum & Bodansky, 1966)). However, the Kms for aspartate and α-ketoglutarate
form distinct clusters corresponding to the mitochondrial and cytoplasmic isozymes
and the values we have reported fall clearly among the cytoplasmic isozymes (p =
0.69) (Figure 4A). We infer from this that the mitochondrial AspAT isozymes are less
concentrated or less active in the samples assayed than the corresponding cytoplasmic
isozymes, consistent with reports for Ascaris lumbricoides (Chu & Chang, 1975). A
similar analysis carried out for the AlaAT isozymes is less clear, but the T.
circumcincta data do appear to be consistent with the cytoplasmic isozymes being at a
lower concentration or less active than the corresponding mitochondrial isozymes (p =
0.75) (Figure 4B). We should point out that χ2 tests of the contingency matrices yield
p < 0.001 for AspAT and p = 0.003 for AlaAT, although the relatively small number of
points prompts us to urge caution. The other commonly used quality measure is
Matthew’s (1975) correlation coefficient, which is related to Cramér’s (1946:282-283)
mean square contingency (φ). All of the quality measures recommended by Vihinen
(2012) indicate that the discriminants shown in Figure 4, A and B, are moderately
useful (Table 1), although we urge caution given the small number of points in the case
of AlaAT (Figure 4B).
The internal consistency of our kinetic parameter estimates can be confirmed
by comparing the equilibrium constant (K′) estimated from the Haldane relation with
the measured value. While such consistency might be considered inevitable, it is not
always the case (Duff et al., 2012). For the ping pong bi bi mechanism characteristic of
aminotransferases (Braunstein, 1973), the relevant Haldane relation is
mBmA
mQmP
r
f
KK
KK
V
VK
2
max
max
=′ ,
where KmA and KmB are the Kms for the substrates (A and B) and KmP and KmQ are those
for the products (P and Q) and the superscripts f and r indicate the forward and reverse
reactions, respectively (Cleland, 1963). Using the parameter estimates for T.
circumcincta AlaAT, the Haldane relation yields K′ = 1.1 ± 0.4, which is similar to the
measured value (adjusted to 30°C) of K′ = 1.29 (Tewari et al., 1998). For AspAT, the
Haldane relation yields K′ = 0.15 ± 0.06, similar to the value at 30°C of K′ = 0.145 ±
0.016 given by Kishore et al. (1998).
Annals of West University of Timişoara, ser. Biology, 2014, vol XVII (2), pp.111-122
119
FIG. 4. Discrimination between mitochondrial (□) and cytoplasmic (○) isozymes of AspAT (A) and AlaAT
(B) using published values of Km for α-ketoglutarate, aspartate and alanine. The dashed curve in each panel
is the boundary between the two isozymes calculated using Fisher’s linear discriminant analysis (using lda
in R) based on those data in the literature for which the isozyme is unambiguously identified (Braunstein,
1973: 427; Burenina, 2001; DeRosa & Swick, 1975; Di Cola et al., 1976; Hopper & Segal, 1962; Huynh et
al., 1980; Kuramitsu et al., 1985; Lowe & Rowe, 1985; Martins et al., 2002; McAllister et al., 2013; Michuda
& Martinez-Carrion, 1969; Nisselbaum & Bodansky, 1966; Orlacchio et al., 1979; Ruscák et al., 1982; Segal
et al., 1962; Swick et al., 1965; Umemura et al., 1994), the values for T. circumcincta (▲) are superimposed
on the plot (but were not used in estimating the curves).
TABLE 1. Quality measures of the linear discriminants shown in Fig. 4.
AspAT AlaAT
Accuracy (%) 88.2 85.7 Negative predictive value (%) 75.0 100.0
Positive predictive value (%) 95.4 76.9
Sensitivity (%) 87.5 100.0 Specificity (%) 90.0 72.7
Matthews correlation coefficient 0.739 0.748
p (χ2) <0.001 0.003
The only previous report of AlaAT activity in T. circumcincta is that of
Walker and Barrett (1991). They reported that Vf/V
r = 0.42 for T. circumcincta,
similar to the values they reported for Nippostrongylus strongyloides (0.50),
Trichostrongylus colubriformis (0.76) and Haemonchus contortus (1.03).
However, Walker and Barrett (1991) used high alanine concentrations in
estimating the rate of alanine deamination (Vf). Such concentrations can be
inhibitory in some species (Bulos & Handler, 1965; Splittstoesser et al., 1976),
although we observed no such substrate inhibition of this reaction by alanine or
glutamate. Nevertheless, the data of Walker and Barrett (1991) might represent
an underestimate of Vf/V
r for T. circumcincta, because we observed that
23.2maxmax
=rf VV , similar to the value of 2.29 for AspAT. However, the internal
consistency of the kinetic parameters we have demonstrated leads us to

MUHAMAD et al: Some properties of aminotransferases from Teladorsagia circumcincta
120
conclude that our estimates are reasonable, despite this apparent inconsistency
with the data of Walker and Barrett (1991) for AlaAT.
We have shown that AspAT and AlaAT have 2.2maxmax ≈rf VV , but AlaAT
has a K′ that is about seven times that of AspAT. We do not know the value of the
mass action ratio (Q) of either enzyme in the cytosol and mitochondria of T.
circumcincta, but if the mitochondrial isoform of AlaAT is the predominant form of
the enzyme (Figure 4B and Table 1), which is characteristic of gluconeogenic tissues
(liver and kidney) in mammals, then the reaction might tend to favour the production
of pyruvate from alanine (DeRosa & Swick, 1975). This implies that QAlaAT < K′AlaAT,
consistent with the depletion of pyruvate in the mitochondrial matrix by the pyruvate
dehydrogenase complex and pyruvate carboxylase. However, Anderson and Garfinkel
(1971) estimated that QAlaAT ≈ K′AlaAT, which implies that AlaAT tends to operate close
to equilibrium and that the reaction can proceed in either direction. In contrast, QAspAT
< K′AspAT (Anderson & Garfinkel, 1971), which prompts us to conclude that the
reaction would tend to favour the formation of glutamate. If this is the case and if the
cytosolic isozyme is the predominant form of AspAT in T. circumcincta (Figure 4A
and Table 1), then this would be consistent with the transfer of reducing equivalents
into the mitochondrial matrix by the malate-aspartate shuttle (Kovacevic, 1972).
CONCLUSIONS
From the kinetic properties of T. circumcincta AlaAT, we infer that the
enzyme is present predominantly as a mitochondrial isozyme favouring pyruvate
formation, consistent with the depletion of pyruvate in the mitochondrial matrix by the
pyruvate dehydrogenase complex and pyruvate carboxylase. In contrast, the kinetic
properties of T. circumcincta AspAT indicate that this enzyme is predominantly a
cytosolic isozyme favouring glutamate formation, which may be consistent with the
transfer of reducing equivalents into the mitochondrial matrix by the malate-aspartate
shuttle.
ACKNOWLEDGEMENTS
This work was funded by Beef + Lamb New Zealand, AGMARDT, Universiti
Malaysia Sarawak and the Public Service Department, Malaysia.
REFERENCES
• Anderson, J. H., Garfinkel, D. 1971. Calculated intracellular distributions of dicarboxylic acids in rat liver. Comput Biomed Res, 4, 43-64.
• Barrett, J. 1991. Amino acid metabolism in helminths. Adv Parasitol, 30, 39-105.
• Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein
utilizing the principle of protein-dye binding. Anal Biochem, 72, 248-254.
• Braunstein, A. E. (1973). Amino group transfer. In The enzymes, vol. 9 (ed. P. D. Boyer), pp. 379-481.

Annals of West University of Timişoara, ser. Biology, 2014, vol XVII (2), pp.111-122
121
• Brown, S., Muhamad, N., Pedley, K. C., Simcock, D. C. 2014. The kinetics of enzyme mixtures. Mol Biol Res Commun, 3, 19-30.
• Bulos, B., Handler, P. 1965. Kinetics of beef heart glutamic-alanine transaminase. J Biol Chem, 240, 3283-
3294.
• Burenina, E. A. 2001. Properties of some enzymes of gluconeogenesis in platyhelminths. J Evol Biochem
Physiol, 37, 111-120.
• Cascante, M., Cortés, A. 1988. Kinetic studies of chicken and turkey liver mitochondrial aspartate
aminotransferase. Biochem J, 250, 805-812.
• Chu, J.-K., Chang, M.-C. 1975. Studies on glutamic pyruvic- and oxaloacetic transaminase of different
organs of Ascaris lumbricoides suis. Korean Journal of Parasitology, 13, 53-59.
• Cleland, W. W. 1963. The kinetics of enzyme-catalyzed reactions with two or more substrates and products. I. Nomenclature and rate equations. Biochim Biophys Acta, 67, 104-137.
• Cramér, H. 1946. Mathematical methods of statistics. Princeton University Press, Princeton.
• Creighton, D. J., Murthy, N. S. R. K. (1990). Stereochemistry of enzyme-catalyzed reactions at carbon. In
The enzymes, vol. 19 (ed. D. S. Sigman and P. D. Boyer), pp. 324-421. Academic Press, Inc., San Diego.
• DeRosa, G., Swick, R. W. 1975. Metabolic implications of the distribution of the alanine aminotransferase
isozymes. J Biol Chem, 250, 7961-7967.
• Di Cola, D., Polidoro, G., Di Ilio, C., Del Boccio, G., Moriggi, M., Politi, L., Scandurra, R. 1976.
Simultaneous purification of mitochondrial and cytoplasmic isozymes of aspartate aminotransferase from beef kidney. Acta Vitaminol Enzymol, 30, 28-37.
• Duff, S. M. G., Rydel, T. J., McClerren, A. L., Zhang, W., Li, J. Y., Sturman, E. J., Halls, C., Chen, S.,
Zeng, J., Peng, J., Kretzler, C. N., Evdokimov, A. 2012. The enzymology of alanine aminotransferse isoforms from Hordeum vulgare and other organisms, and the HvAlaAT crystal structure. Arch Biochem
Biophys, 528, 90-101.
• Grantham, B. D., Barrett, J. 1986. Amino acid catabolism in the nematodes Heligmosomoides polygyrus and
Panagrellus redivivus. 1. Removal of the amino group. Parasitol, 93, 481-493.
• Hopper, S., Segal, H. L. 1962. Kinetic studies of rat liver glutamic-alanine transaminase. J Biol Chem, 237, 3189-3195.
• Huynh, Q. K., Sakakibara, R., Watanabe, T., Wada, H. 1980. Glutamic oxaloacetic transaminase isozymes from rat liver. Purification and physicochemical characterization. J Biochem, 88, 231-239.
• Ihaka, R., Gentleman, R. 1996. R: a language for data analysis and graphics. J Comput Graphical Stat, 5, 299-314.
• Kishore, N., Tewari, Y. B., Goldberg, R. N. 1998. An investigation of the equilibrium of the reaction {L-asparate(aq) + 2-oxoglutarate(aq) = oxaloacetate(aq) + L-glutamate(aq)}. J Chem Thermodynamics, 30,
1373-1384.
• Kovacevic, Z. 1972. Possibility for the transfer of reducing equivalents from the cytosol to the mitochondrial compartment in Ehrlich ascites tumor cells by the malate-aspartate shuttle. Eur J Biochem,
25, 372-378.
• Kuramitsu, S., Inoue, K., Kondo, K., Aki, K., Kagamiyama, H. 1985. Aspartate aminotransferase isozymes
from rabbit liver. Purification and properties. J Biochem, 97, 1337-1345.
• Lowe, P. N., Rowe, A. F. 1985. Aspartate:2-oxoglutarate aminotransferase from Trichomonas vaginalis. Identity of aspartate aminotransferase and aromatic amino acid aminotransferase. Biochem J, 232, 689-695.
• Lowe, P. N., Rowe, A. F. 1986. Aspartate:2-oxoglutarate aminotransferase from Trichomonas vaginalis: comparison with pig heart cytoplasmic enzyme. Comp Biochem Physiol, 85B, 93-99.
• Martinez-Carrion, M., Turano, C., Chiacone, E., Bossa, F., Giartosio, A., Riva, A., Fasella, P. 1967. Isolation and characterization of multiple forms of glutamate-aspartate aminotransferase from pig heart. J
Biol Chem, 242, 2397-2409.
• Martins, M. L. L., Mourato, M. P. F. B., de Varennes e Mendonca, A. P. A. 2002. Characterization of aspartate aminotransferase isoenzymes from leaves of Lupinus albus L. cv Estoril. J Biochem Mol Biol, 35,
220-227.
• Matthews, B. W. 1975. Comparison of the predicted and observed secondary structure of T4 phage
lysozyme. Biochim Biophys Acta, 405, 442-451.
• McAllister, C. H., Facette, M., Holt, A., Good, A. G. 2013. Analysis of the enzymatic properties of a broad
family of alanine aminotransferases. PLoS ONE, 8, e55032.

MUHAMAD et al: Some properties of aminotransferases from Teladorsagia circumcincta
122
• Michuda, C. M., Martinez-Carrion, M. 1969. Distinctions in the equilibrium kinetic constants of the mitochondrial and supernatant isozymes of aspartate transaminase. J Biol Chem, 244, 5920-5927.
• Muhamad, N. (2006). Nitrogen metabolism in Ostertagia (Teladorsagia) circumcincta. PhD thesis, Massey University.
• Muhamad, N., Brown, S., Pedley, K. C., Simpson, H. V. 2005. Some aspects of nitrogen metabolism in Ostertagia circumcincta. N Z J Zool, 32, 278-279.
• Muhamad, N., Simcock, D. C., Pedley, K. C., Simpson, H. V., Brown, S. 2011. The kinetics of glutamate
dehydrogenase of Teladorsagia circumcincta and the lifestyle of the parasite. Comp Biochem Physiol, 159B, 71-77.
• Muhamad, N., Simcock, D. C., Pedley, K. C., Walker, L. R., Simpson, H. V., Brown, S. 2013. Urea output by L3 Teladorsagia circumcincta and some properties of two urea producing enzymes. Iranian J Parasitol,
8, 137-144.
• Muhamad, N., Walker, L. R., Pedley, K. C., Simcock, D. C., Brown, S. 2012. The kinetics of NH3/NH4+
efflux from L3 Teladorsagia circumcincta. Parasitol Int, 61, 487-492.
• Nisselbaum, J. S., Bodansky, O. 1966. Kinetics and electrophoretic properties of the isozymes of aspartate aminotransferase from pig heart. J Biol Chem, 241, 2661-2664.
• Orlacchio, A., Campos-Cavieres, M., Pashev, I., Munn, E. A. 1979. Some kinetic and other properties of the isoenzymes of aspartate aminotransferase isolated from sheep liver. Biochem J, 177, 583-593.
• Ruscák, M., Orlicky, J., Zubor, V., Hager, H. 1982. Alanine aminotransferase in bovine brain: purification and properties. J Neurochem, 39, 210-216.
• Sapan, C. V., Lundblad, R. L., Price, N. C. 1999. Colorimetric protein assay techniques. Biotechnol Appl
Biochem, 29, 99-108.
• Scardi, V., Scotto, P., Iaccarino, M., Scarano, E. 1963. The binding of pyridoxal 5-phosphate to asparatate
aminotransferase of pig heart. Biochem J, 88, 172-175.
• Segal, H. L., Beattie, D. S., Hopper, S. 1962. Purification and properties of liver glutamic-alanine
transaminase from normal and corticoid-treated rats. J Biol Chem, 237, 1914-1920.
• Sharma, R. K., Singh, K. 1977. Studies on glutamic-oxalacetic (GOT) and glutamic-pyruvic (GPT)
transaminases of swine kidney worm Stephanurus dentatus (Diesing, 1839). I. Assay and general properties. Z Parasitenkunde, 54, 251-256.
• Simcock, D. C., Brown, S., Neale, J. D., Przemeck, S. M. C., Simpson, H. V. 2006. L3 and adult Ostertagia
(Teladorsagia) circumcincta exhibit cyanide sensitive oxygen uptake. Exptl Parasitol, 112, 1-7.
• Simcock, D. C., Walker, L. R., Pedley, K. C., Simpson, H. V., Brown, S. 2012. Phosphoenolpyruvate
metabolism in Teladorsagia circumcincta: a critical junction between aerobic and anaerobic metabolism? Exptl Parasitol, 132, 267-273.
• Splittstoesser, W. E., Chu, M. C., Stewart, S. A., Splittstoesser, S. A. 1976. Alanine aminotransferase from Cucurbita moschata cotyledons. Plant Cell Physiol, 17, 83-89.
• Swick, R. W., Barnstein, P. L., Stange, J. L. 1965. The metabolism of mitochondrial proteins. I.
Distribution and characterization of the isozymes of alanine aminotransferase in rat liver. J Biol Chem, 240, 3334-3340.
• Tewari, Y. B., Kishore, N., Goldberg, R. N., Luong, T. N. 1998. An equilibrium and calorimetric study of some transamination reactions. J Chem Thermodynamics, 30, 777-793.
• Umemura, I., Yanagiya, K., Komatsubara, S., Sato, T., Tosa, T. 1994. Purification and some properties of alanine aminotransferase from Candida maltosa. Biosci Biotech Biochem, 58, 283-287.
• Vihinen, M. 2012. How to evaluate performance of prediction methods? Measures and their interpretation
in variation effect analysis. BMC Genomics, 13, S2.
• Walker, J., Barrett, J. 1991. Studies on alanine aminotransferase in nematodes. Int J Parasitol, 21, 377-380.
• Walker, L. R., Simcock, D. C., Neale, J. D., Simpson, H. V., Brown, S. 2007. Thermotolerance of L3 Ostertagia (Teladorsagia) circumcincta and some of its enzymes. Vet Parasitol, 146, 77-82.
Top Related