
PERKEMBANGAN PISANG RAJA NANGKA (Musa sp.) · PDF filepisang Saba dengan eksplan anakan...
6
ISSN 1410-1939 35 PERKEMBANGAN PISANG RAJA NANGKA (Musa sp.) SECARA KULTUR JARINGAN DARI EKSPLAN ANAKAN DAN MERISTEM BUNGA [THE DEVELOPMENT OF BANANA (Musa sp.) CV. RAJA NANGKA VIA TISSUE CULTURE USING SUCKER AND FLORAL MERISTEM EXPLANTS] Rainiyati, Dede Martino, Gusniwati dan Jasminarni Fakultas Pertanian Universitas Jambi Kampus Pinang Masak, Mendalo Darat, Jambi 36361 Abstract An investigation to study the in vitro growth and development of banana cv. Raja Nangka from sucker and floral meristem explants had been conducted at the Plant Biotechnology Laboratory, Agricultural Faculty the University of Jambi from April through to July 2005. This study used completely randomised design, and consisted of two trials: shoot multiplication using BAP (3, 4, and 5 mg L -1 ) in combination with IAA (0, 0.1 and 0.2 mg L -1 ), and root induction using BAP (0, 0.1 and 0.2 mg L -1 ) in combination with IBA (2, 2.5 and 3 mg L - 1 ). Each combination consisted of 10 replicates, and each experimental unit consisted of one explant per culture flask. The results indicated that explant from sucker regenerated shoots within 2 weeks after culture initiation, while explant from floral meristem took longer time to regenerate shoots, i.e. Two months after culture initiation. Key words: in vitro culture, plant biotechnology, plant growth regulator, auxin, cytokinin. PENDAHULUAN Tanaman pisang komersil yang dibudidayakan hingga saat ini adalah triploid (3n) dan tidak mam- pu menghasilkan biji atau partenokarpi, walaupun ada juga yang diploid dan tidak berbiji seperti pi- sang mas. Oleh karena itu pengembangbiakan pi- sang hanya dilakukan secara vegetatif (tanpa per- kawinan) dengan anakan dan kultur jaringan (Sunarjono, 2002). Kultur jaringan merupakan cara pembiakan ve- getatif yang cepat dan secara genetik sifat-sifat ta- naman anak yan gdihasilkan akan sama atau iden- tik dengan induknya. Dalam teknik kultur jaringan yang perlu mendapat perhatian adalah komposisi media kultur dan zat pengatur tumbuh yang tepat serta sumber eksplan yang digunakan untuk meng- hasilkan plantlet di samping faktor lainnya yaitu cahaya, suhu dan kelembaban. Zat pengatur tumbuh mempunyai peran yang sangat penting dalam mengatur pertumbuhan dan perkembangan eksplan di dalam kultur. Pertum- buhan dan morfogenesis eksplan dalam kultur in vitro diatur oleh interaksi dan keseimbangan zat pengatur tumbuh pada media dengan hormon en- dogen yang terdapat dalam eksplan (George dan Sherrington, 1984). Menurut Gunawan (1987) pe- nambahan zat pengatur tumbuh eksogen akan mengubah level zat pengatur tumbuh endogen sel. Perimbangan zat pengatur tumbuh auksin dan sito- kinin yang sesuai akan sangat besar pengaruhnya untuk menghasilkan plantlet. Auksin umumnya berpengaruh terhadap pe- manjangan sel, pembentukan kalus dan akar ad- ventif serta menghambat pembentukan tunas aksi- lar. Dalam konsentrasi rendah auksin akan mema- cu pembentukan akar adventif, sedangkan dalam konsentrasi tinggi mendorong pembentukan kalus (Pierik, 1997). Auksin yang sering dipakai dalam kultur jaringan adalah IAA (Indoleacetic Acid), 2,4-D (2,4-Dichlorophenoxyacetic Acid), IBA (Indolebutyric Acid) dan NAA (Naphtaleneacetic Acid) (George dan Sherrington, 1984). Sitokinin berperan dalam pengaturan pembe- lahan sel dan morfogenesis. Aktivitas utama sito- kinin adalah mendorong pembelahan sel, mengin- duksi pembentukan tunas adventif dan dalam kon- sentrasi tinggi menghambat inisiasi akar (Pierik, 1997). Sitokinin juga menghambat perombakan protein dan klorofil dan menghambat penuaan (senescence) (Wattimena, 1987). Sitokinin yang sering dipakai dalam kultur jaringan adalah BAP (Benzylamino Purine) dan kinetin (George dan Sherrington, 1984). Aplikasi penggunaan auksin dan sitokinin pada tanaman pisang telah banyak dilakukan. Damasco dan Barba (1984) melakukan penelitian terhadap pisang Saba dengan eksplan anakan diperoleh se- kitar 200.000 bibit dalam waktu 10 bulan. Zat pengatur tumbuh yang digunakan adalah 10 mg L -1
Transcript of PERKEMBANGAN PISANG RAJA NANGKA (Musa sp.) · PDF filepisang Saba dengan eksplan anakan...

ISSN 1410-1939
35
PERKEMBANGAN PISANG RAJA NANGKA (Musa sp.) SECARA KULTUR
JARINGAN DARI EKSPLAN ANAKAN DAN MERISTEM BUNGA
[THE DEVELOPMENT OF BANANA (Musa sp.) CV. RAJA NANGKA VIA
TISSUE CULTURE USING SUCKER AND FLORAL MERISTEM EXPLANTS]
Rainiyati, Dede Martino, Gusniwati dan Jasminarni
Fakultas Pertanian Universitas Jambi
Kampus Pinang Masak, Mendalo Darat, Jambi 36361
Abstract
An investigation to study the in vitro growth and development of banana cv. Raja Nangka from sucker and
floral meristem explants had been conducted at the Plant Biotechnology Laboratory, Agricultural Faculty the
University of Jambi from April through to July 2005. This study used completely randomised design, and
consisted of two trials: shoot multiplication using BAP (3, 4, and 5 mg L-1) in combination with IAA (0, 0.1 and
0.2 mg L-1), and root induction using BAP (0, 0.1 and 0.2 mg L-1) in combination with IBA (2, 2.5 and 3 mg L-
1). Each combination consisted of 10 replicates, and each experimental unit consisted of one explant per
culture flask. The results indicated that explant from sucker regenerated shoots within 2 weeks after culture
initiation, while explant from floral meristem took longer time to regenerate shoots, i.e. Two months after
culture initiation.
Key words: in vitro culture, plant biotechnology, plant growth regulator, auxin, cytokinin.
PENDAHULUAN
Tanaman pisang komersil yang dibudidayakan
hingga saat ini adalah triploid (3n) dan tidak mam-
pu menghasilkan biji atau partenokarpi, walaupun
ada juga yang diploid dan tidak berbiji seperti pi-
sang mas. Oleh karena itu pengembangbiakan pi-
sang hanya dilakukan secara vegetatif (tanpa per-
kawinan) dengan anakan dan kultur jaringan
(Sunarjono, 2002).
Kultur jaringan merupakan cara pembiakan ve-
getatif yang cepat dan secara genetik sifat-sifat ta-
naman anak yan gdihasilkan akan sama atau iden-
tik dengan induknya. Dalam teknik kultur jaringan
yang perlu mendapat perhatian adalah komposisi
media kultur dan zat pengatur tumbuh yang tepat
serta sumber eksplan yang digunakan untuk meng-
hasilkan plantlet di samping faktor lainnya yaitu
cahaya, suhu dan kelembaban.
Zat pengatur tumbuh mempunyai peran yang
sangat penting dalam mengatur pertumbuhan dan
perkembangan eksplan di dalam kultur. Pertum-
buhan dan morfogenesis eksplan dalam kultur in
vitro diatur oleh interaksi dan keseimbangan zat
pengatur tumbuh pada media dengan hormon en-
dogen yang terdapat dalam eksplan (George dan
Sherrington, 1984). Menurut Gunawan (1987) pe-
nambahan zat pengatur tumbuh eksogen akan
mengubah level zat pengatur tumbuh endogen sel.
Perimbangan zat pengatur tumbuh auksin dan sito-
kinin yang sesuai akan sangat besar pengaruhnya
untuk menghasilkan plantlet.
Auksin umumnya berpengaruh terhadap pe-
manjangan sel, pembentukan kalus dan akar ad-
ventif serta menghambat pembentukan tunas aksi-
lar. Dalam konsentrasi rendah auksin akan mema-
cu pembentukan akar adventif, sedangkan dalam
konsentrasi tinggi mendorong pembentukan kalus
(Pierik, 1997). Auksin yang sering dipakai dalam
kultur jaringan adalah IAA (Indoleacetic Acid),
2,4-D (2,4-Dichlorophenoxyacetic Acid), IBA
(Indolebutyric Acid) dan NAA (Naphtaleneacetic
Acid) (George dan Sherrington, 1984).
Sitokinin berperan dalam pengaturan pembe-
lahan sel dan morfogenesis. Aktivitas utama sito-
kinin adalah mendorong pembelahan sel, mengin-
duksi pembentukan tunas adventif dan dalam kon-
sentrasi tinggi menghambat inisiasi akar (Pierik,
1997). Sitokinin juga menghambat perombakan
protein dan klorofil dan menghambat penuaan
(senescence) (Wattimena, 1987). Sitokinin yang
sering dipakai dalam kultur jaringan adalah BAP
(Benzylamino Purine) dan kinetin (George dan
Sherrington, 1984).
Aplikasi penggunaan auksin dan sitokinin pada
tanaman pisang telah banyak dilakukan. Damasco
dan Barba (1984) melakukan penelitian terhadap
pisang Saba dengan eksplan anakan diperoleh se-
kitar 200.000 bibit dalam waktu 10 bulan. Zat
pengatur tumbuh yang digunakan adalah 10 mg L-1

Jurnal Agronomi Vol. 11 No. 1, Januari – Juni 2007
36
BAP dan 10 mg L-1
kinetin. Selanjutnya Hwang
(1984) memperoleh 5 - 10 tunas pisang Cavendish
dalam waktu 6 - 8 minggu pada media dengan 2
mg L-1
IAA + 2 mg L-1
kinetin dan 160 mg L-1
adenin sulfat.
Respon dari beberapa jenis pisang pada media
yang sama memberikan hasil yang berbeda-beda
(Damasco dan Barba, 1984; Banerjee dan de
Langhe, 1985). Vuylsteke dan de Langhe (1984)
menyatakan tipe genom pisang mempengaruhi laju
perbanyakan. Dari empat genom yang ditanam pa-
da media MS yang mengandung 10 mg L-1
BAP,
daya multiplikasi tertinggi terjadi pada genom
ABB kemudian diikuti AAB, AAA dan AA. Hasil
penelitian Sukma (1994) menunjukkan daya multi-
plikasi pisang tertinggi diperoleh pada pisang Am-
bon (AAA), diikuti oleh pisang barangan (AAA),
pisang raja bulu (AAB dan pisang mas (AA) pada
media yang mengandung 10,5 mg L-1
BAP + 3 mg
L-1
IAA.
Penelitian yang dilakukan Rubbyanto (1992)
pada pisang mas menunjukkan bahwa perangsang-
an multiplikasi lebih baik pada perlakuan BAP
5mg L-1
+ IBA 1 mg L-1
dengan menghasilkan 5,2
tunas dalam waktu 5 minggu. Wijayanti (1995)
mendapatkan 4,4 tunas pisang Ambon dalam wak-
tu 8 minggu pada perlakukan 10 mg L-1
BAP dan
perlakuan 5 mg L-1
BAP + 5 mg L-1
2-IP, di mana
masing-masing perlakuan ditambahkan 0 mg L-1
NAA.
Kebutuhan zat pengatur tumbuh bagi pertum-
buhan dan perkembangan tanaman pisang sangat
tergantung dari sifat genetik dan tingkat konsentra-
si auksin dan sitokinin yang diberikan. Jenis pisang
yang berbeda akan memberikan respon pertum-
buhan yang berbeda tanggapannya terhadap kese-
imbangan auksin dan sitokinin yang diberikan pa-
da media. Untuk itu perlu dicari metoda perba-
nyakan pisang raja nangka yang efektif dengan ca-
ra mencoba kombinasi beberapa konsentrasi auk-
sin dan sitokinin berdasarkan hasil penelitian yang
sudah ada.
Hasil yang diharapkan dari percobaan ini ada-
lah didapatkan metoda perbanyakan pisang raja
nangka yang efektif dengan ditemukannya kombi-
nasi auksin dan sitokinin yang terbaik untuk meng-
hasilkan plantlet pisang raja nangka yang berasal
dari dua sumber eksplan anakan dan meristem bu-
nga.
BAHAN DAN METODA
Penelitian ini dilakukan di Laboratorium Bio-
teknologi Tanaman Fakultas Pertanian Universitas
Jambi selama 7 bulan, mulai bulan April sampai
dengan bulan Oktober 2005.
Bahan tanaman (eksplan) yang digunakan ada-
lah tunas in vitro pisang Raja Nangka yang diini-
siasi dari anakan muda yang sehat dan segar de-
ngan tinggi 5 - 20 cm dan meristem bunga pisang.
Tunas ditanam berukuran lebih-kurang 1,5 cm. Se-
tiap botol kultur berisi 1 eksplan.
Bahan kimia yang digunakan adalah bahan da-
sar media MS yang dimodifikasi dan bahan-bahan
tambahan. Zat pengatur tumbuh yang digunakan
adalah BAP, kinetin, NAA dan IAA yang konsen-
trasinya sesuai dengan perlakuan. Sebagai bahan
pemadat digunakan agar 7 g L-1
, sedangkan untuk
sterilisasi alat digunakan alkohol 95%.
Alat-alat yang digunakan adalah peralatan un-
tuk pembuatan media, yaitu timbangan analitik, pi-
pet, labu takar, pengaduk gelas (sudip), pH meter
dan otoklaf. Untuk tempat media digunakan botol
biakan berukuran 100-mL dengan penutup alumi-
nium foil. Peralatan yang digunakan pada saat pe-
nanaman adalah kotak pindah (laminar air flow
cabinet), petridish, alat diseksi (pinset, skalpel dan
gunting) hand sprayer, lampu spiritus, rak kultur
dan lampu neon.
Penelitian ini terdiri dari 2 tahap percobaan.
Percobaan pertama bertujuan untuk multiplikasi
tunas, sedangkan percobaan kedua bertujuan untuk
perakaran. Percobaan pertama adalah percobaan
faktorial yang disusun dalam Rancangan Acak
Lengkap yang terdiri dari tiga faktor. Faktor perta-
ma adalah sumber eksplan: anakan 1 dan anakan 2,
faktor ke-dua adalah beberapa konsentrasi BAP (3,
4 dan 5 mg L-1
) dan faktor ketiga adalah beberapa
konsentrasi IAA (0, 0,1 dan 0,2 mg L-1
). Percobaan
tahap ke-dua adalah lanjutan tahap pertama juga
percobaan faktorial yang disusun dalam Rancang-
an Acak Lengkap yang terdiri dari tiga faktor. Fak-
tor pertama adalah sumber eksplan: anakan 1 dan
anakan 2, faktor ke-dua adalah beberapa konsen-
trasi BAP (0, 0,1, 0.2 mg L-1
) dan faktor ketiga
adalah beberapa konsentrasi IBA (2, 2,5 dan 3 mg
L-1
). Masing-masing set percobaan terdiri dari 24
kombinasi perlakuan dengan masing-masing unit
percobaan ini terdiri dari 1 eksplan per botol di
mana setiap unit percobaan dilakukan 10 ulangan.
Peubah yang diamati dalam 2 tahap percobaan
ini adalah persentase tunas yang terbentuk, persen-
tase tunas yang berakar, jumlah tunas, jumlah daun
dan jumlah akar. Pengamatan dilakukan dengan
cara menghitung jumlah tunas, daun dan akar yang
terbentuk setiap minggu. Data yang dikumpulkan
selama penelitian berlangsung dianalisis menggu-
nakan sidik ragam dan dilanjutkan dengan uji Dun-
can (DNMRT) pada taraf α = 5%.
Rainiyati et al.: Kultur Jaringan Pisang Raja Nangka.
37
HASIL DAN PEMBAHASAN
Hasil penelitian menunjukkan bahwa eksplan
menghasilkan tunas dua minggu setelah ditanam
pada media perlakuan. Tunas yang terbentuk akan
terus bertambah baik ukuran maupun jumlahnya.
Menurut Taji et al. (2002) bilamana pertumbuhan
diukur secara terus-menerus berdasarkan waktu,
maka akan diperoleh kurva berbentuk huruf S.
Bentuk kurva pertumbuhan demikian dapat diper-
oleh dari pertumbuhan sel-sel tunggal, organ ta-
naman atau keseluruhan tanaman. Pada penelitian
ini pembentukan tunas pada masing-masing eks-
plan berbeda-beda, baik dilihat dari waktu terben-
tuknya dan jumlah tunas yang terbentuk. Adanya
perbedaan ini disebabkan karena adanya perbedaan
konsentrasi dari masing-masing kombinasi zat
pengatur tumbuh yang diberikan. Untuk lebih jelas
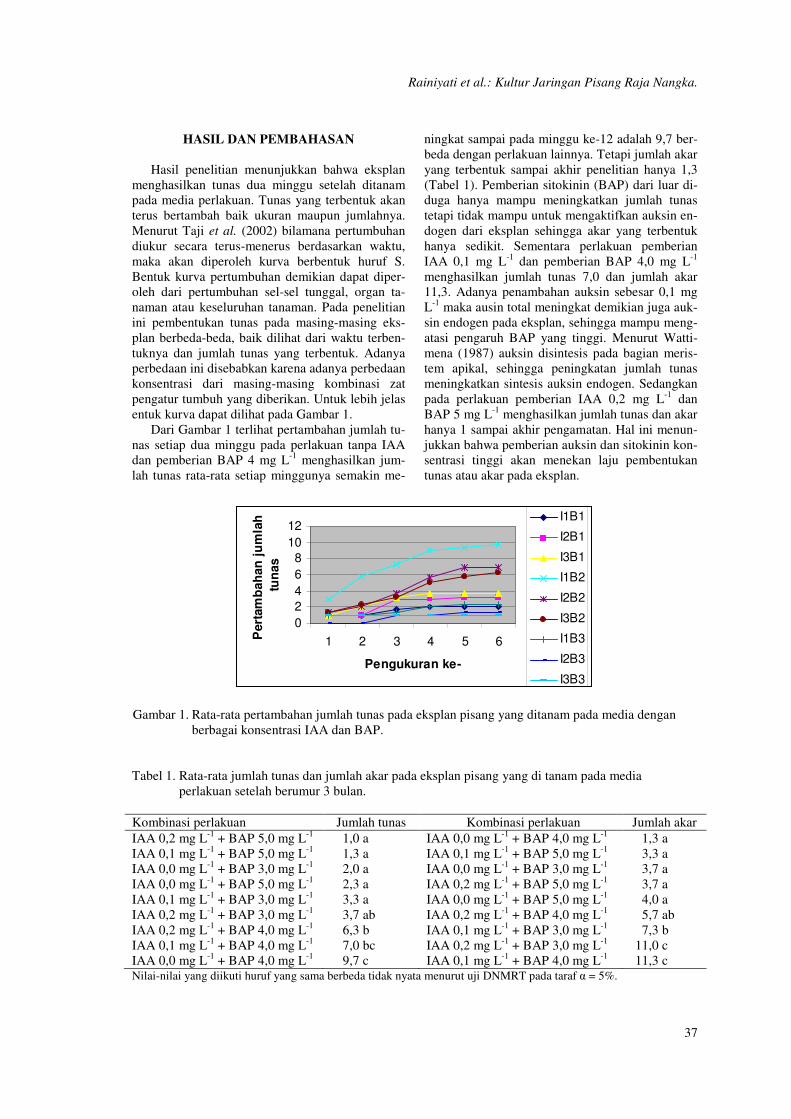
entuk kurva dapat dilihat pada Gambar 1.
Dari Gambar 1 terlihat pertambahan jumlah tu-
nas setiap dua minggu pada perlakuan tanpa IAA
dan pemberian BAP 4 mg L-1
menghasilkan jum-
lah tunas rata-rata setiap minggunya semakin me-
ningkat sampai pada minggu ke-12 adalah 9,7 ber-
beda dengan perlakuan lainnya. Tetapi jumlah akar
yang terbentuk sampai akhir penelitian hanya 1,3
(Tabel 1). Pemberian sitokinin (BAP) dari luar di-
duga hanya mampu meningkatkan jumlah tunas
tetapi tidak mampu untuk mengaktifkan auksin en-
dogen dari eksplan sehingga akar yang terbentuk
hanya sedikit. Sementara perlakuan pemberian
IAA 0,1 mg L-1
dan pemberian BAP 4,0 mg L-1
menghasilkan jumlah tunas 7,0 dan jumlah akar
11,3. Adanya penambahan auksin sebesar 0,1 mg
L-1
maka ausin total meningkat demikian juga auk-
sin endogen pada eksplan, sehingga mampu meng-
atasi pengaruh BAP yang tinggi. Menurut Watti-
mena (1987) auksin disintesis pada bagian meris-
tem apikal, sehingga peningkatan jumlah tunas
meningkatkan sintesis auksin endogen. Sedangkan
pada perlakuan pemberian IAA 0,2 mg L-1
dan
BAP 5 mg L-1
menghasilkan jumlah tunas dan akar
hanya 1 sampai akhir pengamatan. Hal ini menun-
jukkan bahwa pemberian auksin dan sitokinin kon-
sentrasi tinggi akan menekan laju pembentukan
tunas atau akar pada eksplan.
0
2
4
6
8
10
12
1 2 3 4 5 6
Pengukuran ke-
Pert
am
bah
an
ju
mla
h
tun
as
I1B1
I2B1
I3B1
I1B2
I2B2
I3B2
I1B3
I2B3
I3B3
Gambar 1. Rata-rata pertambahan jumlah tunas pada eksplan pisang yang ditanam pada media dengan
berbagai konsentrasi IAA dan BAP.
Tabel 1. Rata-rata jumlah tunas dan jumlah akar pada eksplan pisang yang di tanam pada media
perlakuan setelah berumur 3 bulan.
Kombinasi perlakuan Jumlah tunas Kombinasi perlakuan Jumlah akar
IAA 0,2 mg L-1
+ BAP 5,0 mg L-1
1,0 a IAA 0,0 mg L-1
+ BAP 4,0 mg L-1
1,3 a
IAA 0,1 mg L-1
+ BAP 5,0 mg L-1
1,3 a IAA 0,1 mg L-1
+ BAP 5,0 mg L-1
3,3 a
IAA 0,0 mg L-1
+ BAP 3,0 mg L-1
2,0 a IAA 0,0 mg L-1
+ BAP 3,0 mg L-1
3,7 a
IAA 0,0 mg L-1
+ BAP 5,0 mg L-1
2,3 a IAA 0,2 mg L-1
+ BAP 5,0 mg L-1
3,7 a
IAA 0,1 mg L-1
+ BAP 3,0 mg L-1
3,3 a IAA 0,0 mg L-1
+ BAP 5,0 mg L-1
4,0 a
IAA 0,2 mg L-1
+ BAP 3,0 mg L-1
3,7 ab IAA 0,2 mg L-1
+ BAP 4,0 mg L-1
5,7 ab
IAA 0,2 mg L-1
+ BAP 4,0 mg L-1
6,3 b IAA 0,1 mg L-1
+ BAP 3,0 mg L-1
7,3 b
IAA 0,1 mg L-1
+ BAP 4,0 mg L-1
7,0 bc IAA 0,2 mg L-1
+ BAP 3,0 mg L-1
11,0 c
IAA 0,0 mg L-1
+ BAP 4,0 mg L-1
9,7 c IAA 0,1 mg L-1
+ BAP 4,0 mg L-1
11,3 c Nilai-nilai yang diikuti huruf yang sama berbeda tidak nyata menurut uji DNMRT pada taraf α = 5%.

Jurnal Agronomi 11(1):
38
Rata-rata jumlah tunas yang terbentuk pada
kombinasi perlakuan tanpa IAA dan pemberian
BAP 4 mg L-1
dua minggu setelah tanam adalah 3
tunas sedangkan pada perlakuan lainnya 1 - 1,3 tu-
nas. Pertambahan jumlah tunas semakin meningkat
setiap minggunya sehingga terbentuk multiplikasi
tunas setelah minggu ke-12. Hal ini menunjukkan
bahwa pemberian sitokinin dengan konsentrasi 4
mg L-1
pada media sudah cukup untuk pembentuk-
an multiplikasi tunas namun akar tidak terbentuk.
Sitokinin biasanya tidak digunakan pada tahap
perakaran karena aktifitasnya dapat menghambat
pembentukan akar dan menghalangi pertumbuhan
akar, serta menghambat pengaruh auksin terhadap
inisiasi akar pada kultur jaringan sejumlah spesies
tertentu (George dan Sherrington, 1984). Menurut
Bhojwani dan Rhazdan (1983), sitokinin (BAP) sa-
ngat efektif dalam merangsang pembentukan tu-
nas. Semakin tinggi konsentrasi sitokinin, jumlah
tunas yang terbentuk semakin bertambah tetapi
pembentukan masing-masing tunas terhambat. Pa-
da penelitian ini peningkatan jumlah sitokinin
menjadi 5 mg L-1
setelah minggu ke-12 tidak terja-
di penambahan jumlah tunas. Perkembangan jum-
lah tunas pada perlakuan tanpa IAA dan BAP 4 mg
L-1
dari umur 1 minggu setelah tanam sampai ter-
bentuk multiplikasi tunas seperti yang terlihat pada
Gambar 2.
Multiplikasi tunas mulai terbentuk setelah dua
minggu tanaman dipindahkan ke media pertunas-
an. Multiplikasi tunas terbentuk karena beberapa
faktor yang mendukung yaitu eksplan dan ling-
kungan tumbuh (suhu, cahaya dan zat pengatur
tumbuh). Sedangkan nodul yang terbentuk dari
eksplan bunga akan memperlihatkan terjadinya
multiplikasi nodul setelah 4 bulan pada medium
pertunasan. Tunas yang telah terbentuk dari eks-
plan anakan setelah berumur 2 bulan dipindahkan
ke media perakaran untuk membentuk plantlet.
Perakaran terbentuk setelah 2 minggu tunas dipin-
dahkan ke media perakaran dan dipelihara sampai
plantlet siap untuk diaklimatisasi. Pembentukan
akar ini dipengaruhi oleh pemberian zat pengatur
tumbuh auksin dengan konsentrasi tinggi yaitu
IBA 2 mg L-1
dan sitokinin dengan konsentrasi
rendah yaitu 0,5 mg L-1
BAP. Adanya perimbang-
an zat pengatur tumbuh ini pada media menghasil-
kan sistem perakaran yang baik pada kultur yang
berasal dari anakan
Untuk melihat lebih jauh pengaruh kombinasi
perlakuan terhadap jumlah tunas dan akar dapat
terlihat pada Tabel 1. Di sini jelas terlihat bahwa
perimbangan pemberian auksin dan sitokinin pada
eksplan pisang sangatlah menentukan. Kecende-
rungan pemberian auksin yang tinggi (IAA 0,2 mg
L-1) dan sitokinin yang rendah (BAP 3 mg
L-1) eks-
plan akan menghasilkan tunas sedikit dan akar
yang banyak, namun peningkatan sitokinin yang
tinggi 5 mg L-1
bahkan menekan pembentukan tu-
nas. Pada penelitian ini kombinasi perlakuan yang
terbaik untuk menghasilkan plantlet pisang adalah
pemberian IAA 0,1 mg L-1
+ BAP 4 mg L-1
yang
menghasilkan jumlah tunas 7 dan jumlah akar
11,3.
Pemberian BA dengan konsentrasi tinggi (5 mg L-1
) pada media memacu sitokinin endogen dari
eksplan dalam merangsang pembentukan tunas dan
memacu pembentukan multiplikasi tunas. Menurut
Bhojwani dan Rhazdan (1983), sitokinin (BAP) sa-
ngat efektif dalam merangsang pembentukan tu-
nas, semakin tinggi konsentrasi sitokinin jumlah
tunas yang terbentuk semakin bertambah tetapi
pembentukan masing-masing tunas terhambat.
Pemberian konsentrasi BAP tinggi ini mengham-
bat terbentuknya akar. Terbentuknya akar maka
pada media perakaran diberikan auksin (IBA) de-
ngan konsentrasi tinggi 2,5 mg L-1
, sehingga jum-
lah auksin total eksplan meningkat dengan penam-
bahan tersebut. Auksin endogen hasil sintesis tunas
juga meningkat sehingga mampu mengatasi penga-
ruh BAP. Wattimena (1987) menyatakan auksin
disintesis pada bagian meristem apikal, sehingga
peningkatan jumlah tunas meningkatkan sintesis
auksin endogen. Pengaruh kombinasi perlakuan
pada penelitian ini dapat dilihat pada Gambar 3.
Gambar 2. Tunas yang terbentuk dari anakan umur
1 minggu (A), 2 minggu (B) dan 3 bu-
lan (C) setelah dipindahkan ke media
perlakuan.
Eksplan yang berasal dari bunga pisang pada
awalnya membentuk nodul-nodul yang embrioge-
nik, setelah 6 bulan pada media perakaran terjadi
multiplikasi nodul, pembentukan tunas yang kemu-
dian diikuti pembentukan akar (Gambar 4). Menu-
rut Wattimena et al. (1992). Nodul merupakan se-
kelompok sel pada tempat tertentu dalam kalus
yang menyerupai kambium, yang sering juga dise-
but meristemoid. Hal ini menungkinkan sel aktif
membelah. Multiplikasi diduga berasal dari sel
peri-peri yang membelah membentuk nodul baru,
namun secara pasti belum dipelajari lebih jauh asal
multiplikasi.

Rainiyati et al.: Kultur Jaringan Pisang Raja Nangka.
39
Tunas tersebut muncul dari nodul-nodul yang
embriogenik. Lamanya waktu yang dibutuhkan un-
tuk membentuk tunas pada nodul tersebut diduga
karena perimbangan zat pengatur tumbuh yang ada
pada media belum tepat sehingga belum mampu
untuk memacu pertumbuhan tunas dengan cepat.
Gambar 3. Tunas yang terbentuk dari anakan 2 bu-
lan setelah dipindahkan ke media per-
lakuan.
Gambar 4. Tunas yang terbentuk pada eksplan bu-
nga pisang setelah 4 bulan pada medi-
um pertunasan (A) dan 6 bulan (B) pa-
da medium pengakaran.
KESIMPULAN
1. Eksplan yang berasal dari anakan membentuk
tunas setelah 2 minggu dipindahkan ke media
MS, sedangkan eksplan yang berasal dari me-
ristem bunga pisang tidak langsung menghasil-
kan tunas tetapi membentuk nodul-nodul yang
embriogenik
2. Multiplikasi tunas pada eksplan yang berasal
dari anakan akan terjadi setelah tanaman 2 bu-
lan dipindahkan ke media pertunasan. Multi-
plikasi nodul pada eksplan yang berasal dari
bunga terjadi 4 bulan setelah dipindahkan ke
media pertunasan, sedangkan tunas baru mun-
cul setelah tanaman berumur 6 bulan.
DAFTAR PUSTAKA
Banerjee, N. dan E. de Langhe. 1985. A tissue culture
technique for rapid clonal propagation and storage
under minimal growth condition of Musa (banana
and plantain). Plant Cell Reports 4: 351-354.
Bhojwani, S. S. dan M. K. Razdan. 1983. Plant Tissue
Culture: Theory and Practice. Development in Crop
Science 5. Elsevier Press, Amsterdam.
Damasco, O. P. dan R. C. Barba. 1984. In vitro culture
of Saba Banana (Musa sp. cv. Saba (BBB)).
Philosophy of Agriculture 67: 351-358.
George, E. F. dan P. D. Sherrington. 1984. Plant
Propagation by Tissue Culture. Exegetics Limited,
England.
Gunawan, L. W. 1987. Teknik kultur jaringan. Pusat
Antar Universitas Institut Pertanian Bogor, Bogor.
Hwang, S. C. 1984. Cultivation of banana using plantlet
from meristem culture. Horticultural Science 19:
231-233.
Pierik, R. L. M. 1997. In Vitro Culture of Higher Plants.
Kluwer Academic Publishers, Dordrecht, The
Netherlands.
Rubbyanto. 1992. Pengaruh Beberapa Macam Sitokinin
terhadap Pertumbuhan Pucuk Pisang Mas dalam
Kultur In Vitro. Skripsi Sarjana. Fakultas Pertanian
Institut Pertanian Bogor, Bogor.
Sukma, D. 1994. Pengaruh IAA dan BAP terhadap
Perbanyakan Tunas Mikro Pisang Mas (Musa
accuminata L. AA Group), Ambon dan Barangan
(Musa accuminata L. AAA Group), dan Raja Bulu
(Musa accuminata L. AAB Group) secara in vitro.
Skripsi Sarjana. Fakultas Pertanian Institut Pertanian
Bogor, Bogor.
Sunarjono, H. 2002. Budidaya Pisang dengan Bibit
Kultur Jaringan. Penebar Swadaya, Jakarta.
Taji, A., P. Kumar dan P. Lakshmanan. 2002. In Vitro
Plant Breeding. Haworth Press, Inc., New York.
Vuylsteke, D. dan E. de Langhe. 1984. Feasibility of in
vitro propagation of bananas and plantains. Tropical
Agriculture 62: 323-328.
Wattimena, G. A. 1987. Zat Pengatur Tumbuh Tanaman.
Pusat Antar Universitas Bioteknologi Institut
Pertanian Bogor, Bogor.
Wattimena, G. A., L. W. Gunawan, N. A. Mattjik, E.
Sjamsudin, N. M. A. Wiendi dan Ernawati. 1992.
Bioteknologi Tanaman. Institut Pertanian Bogor,
Bogor.
Wijayanti, N. 1995. Pengaruh Kombinasi BAP dan 2-iP
terhadap Multiplikasi Tunas Pisang Ambon Kuning
[Musa acuminata (AAA grup)] melalui Kultur In
Vitro. Skripsi Sarjana. Fakultas Pertanian Institut
Pertanian Bogor, Bogor.

Jurnal Agronomi Vol. 11 No. 1, Januari – Juni 2007
40


















